Neuromuscular junctions were generated by co-culturing optogenetic hiPSC-derived motoneurons with non-optogenetic skeletal muscle tissue. Human primary skeletal myoblasts (SkM) were seeded into the platforms and differentiated into multinucleated myotubes using the 2-week protocol. The optogenetic motoneurons were differentiated separately, but in parallel with the myotube differentiation, and then seeded into the platform (Figure 1). The tissues began contracting in response to blue light stimulation 7-12 days after MN seeding, indicating successful development and maturation of the NMJs. The NMJs can be cultured, stimulated, and imaged for up to 40 days after motoneuron seeding, but the optimal time point occurs on day 1619. The morphological and biochemical validation of the differentiations and presence of the optogenetic proteins are shown in Supplementary Figure 1 and have been validated in previous studies18,19.
To demonstrate that the protocol can be easily adapted to different culture systems, the strategy was tested using two different compartmentalized reactors: a microfluidic device and an open-well bioreactor. Both systems have a similar design, with one chamber provided with pillars for the generation of skeletal muscle tissues and a second chamber for the culture of 3D motoneuron aggregates that can extend axons to innervate the skeletal tissues (Figure 2A,B). However, the two systems have different scales, with the microfluidic device yielding 1 mm long skeletal muscle tissues comprising approximately 20,000 myoblasts (Figure 2C) and the open-well bioreactor system resulting in 4 mm long muscle tissues comprising approximately 450,000 myoblasts (Figure 2D).
To allow for controlled stimulation of the optogenetic NMJs, an optical setup was built, comprising of a red 637 nm LED for brightfield illumination, a blue 488 nm LED for activation of ChR2-MNs, and a blue light filter between the tissue sample and objective to prevent blue light from interfering with the image analysis (Figure 3A). The LEDs were powered by LED drivers and controlled precisely via an Arduino microprocessor. NMJ function was evaluated by simultaneously acquiring time-lapse videos while stimulating MNs with blue light using a custom microscope macro-script paired with the Arduino code (Figure 3B). The protocol increased the frequency of stimulation from 0.2 Hz to 2 Hz in 30 steps. Once the movies were acquired, the tissue function was analyzed using a custom MATLAB package (Figure 3C). The analysis measured tissue displacement, generated a contractility trace, and synchronized it with the light stimulation protocol. It then determined the fraction of effective pulses (F) and an expected fraction of effective pulses (E) to account for potential unstimulated contractions. The expected fraction (E) was calculated as the fraction of pulses that would be labeled as effective if the contractions were randomly distributed within the 30 s. The tissue score was then calculated as (F-E)/(1-E), with a value between 0 and 1 (Figure 3C). The code determines if a contraction is triggered based on the time between a light pulse and the start of the contractility peak, as shown in Figure 3D. This automated, unbiased method allows for repeated characterization of NMJ function in large sample sizes and over time. The code includes tunable parameters that can be adjusted after processing to ensure correct scoring (Supplementary Figure 2).
The system's ability to quantitatively characterize NMJ function over time was demonstrated by imaging a group of tissues for 11 days (day 9 to day 20 after motoneuron seeding). The system captured improvement of NMJ function in tissues, showing an increase in triggered responses 1 week after tissue fabrication (Figure 4A), followed by a stable function for an additional week (Figure 4B). To verify that muscle stimulation occurred through the NMJs, another group of tissues was analyzed before and after 20 min of incubation with 5 μg/mL of the neurotoxin α-bungarotoxin (BTX), which binds specifically and irreversibly to acetylcholine receptors. BTX stopped all triggered and spontaneous contractions, resulted in a complete disruption of NMJ function, and demonstrated that light stimulation of the tissues requires a functional NMJ (Figure 4C,D). Serial dilutions of BTX were initially tested to identify the optimal BTX concentration (results not shown).
To evaluate the translational capability of the system, NMJ function was assessed before and after the addition of serum from five myasthenia gravis patients. Analysis of the tissues treated with 20% MG serum for 48 h showed decreased function compared to control tissues treated with serum from healthy donors (Figure 4E,F). The engineered NMJs were also evaluated again 48 h after removal and washout of the serum and showed recovery of function (Figure 4F). Additionally, increasing concentrations of patient serum and normal human serum control (5%, 20%, 40%) were tested to identify the optimal concentration that affected tissue function. The results showed the ability of the system to characterize and quantify NMJ function and to model myasthenia gravis in vitro.
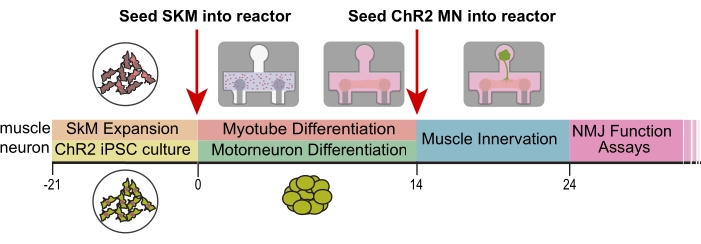
Figure 1. Experimental design and timeline. Timeline (days) of differentiation and seeding into platforms. SkM = skeletal muscle, ChR2 = optogenetic motoneurons, NMJ = neuromuscular junction. This figure has been modified with permission from Vila et al.18. Please click here to view a larger version of this figure.
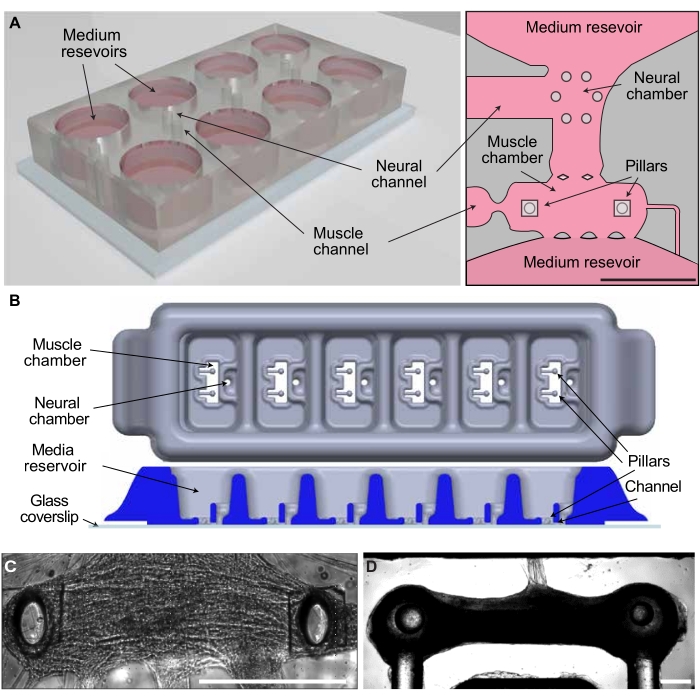
Figure 2. Co-culture systems for the generation of 3D NMJs. (A) Schematic of the microfluidic platform. (B) Schematic of the open-well PDMS bioreactor. (C) Innervated skeletal muscle tissue in the microfluidic platform. (D) Innervated tissue in the open-well bioreactor. Scale bar 0.5 mm. This figure has been modified with permission from Vila et al.18. Please click here to view a larger version of this figure.
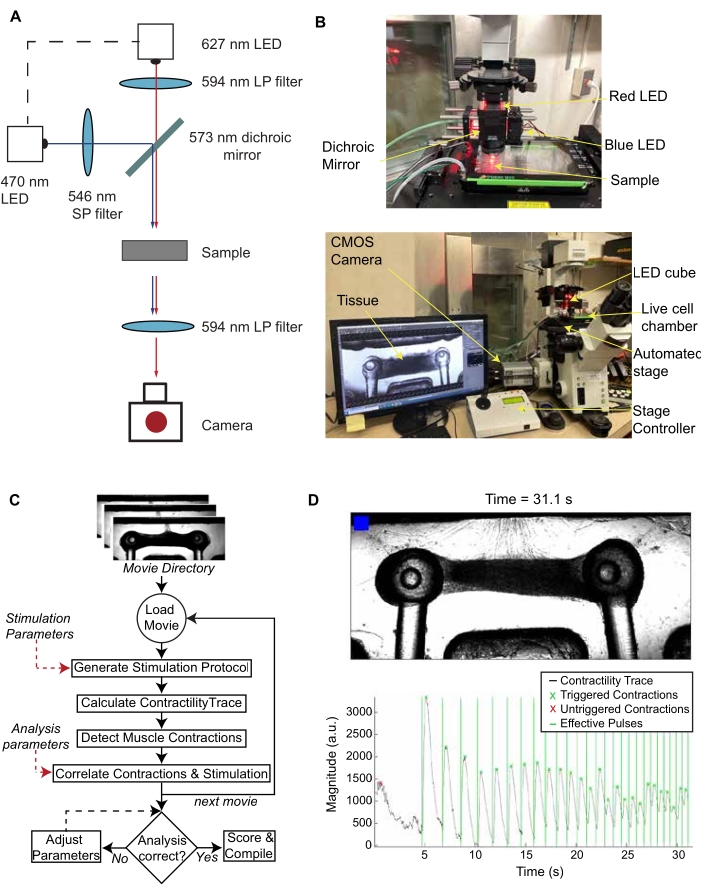
Figure 3. Stimulation, recording, and analysis of NMJ function. (A) Schematic of LED set up and light path. (B) Optical set-up with an inverted microscope, live cell chamber, automated stage, and sCMOS camera. (C) Algorithm for the batch processing of optical stimulation videos of NMJ cultures. (D) Representative frame of the video generated by the MATLAB code and the corresponding contractility trace graph. The blinking blue box in the video and the vertical lines in the graph indicate when the blue light stimulation occurs. This figure has been modified with permission from Vila et al.18. Please click here to view a larger version of this figure.
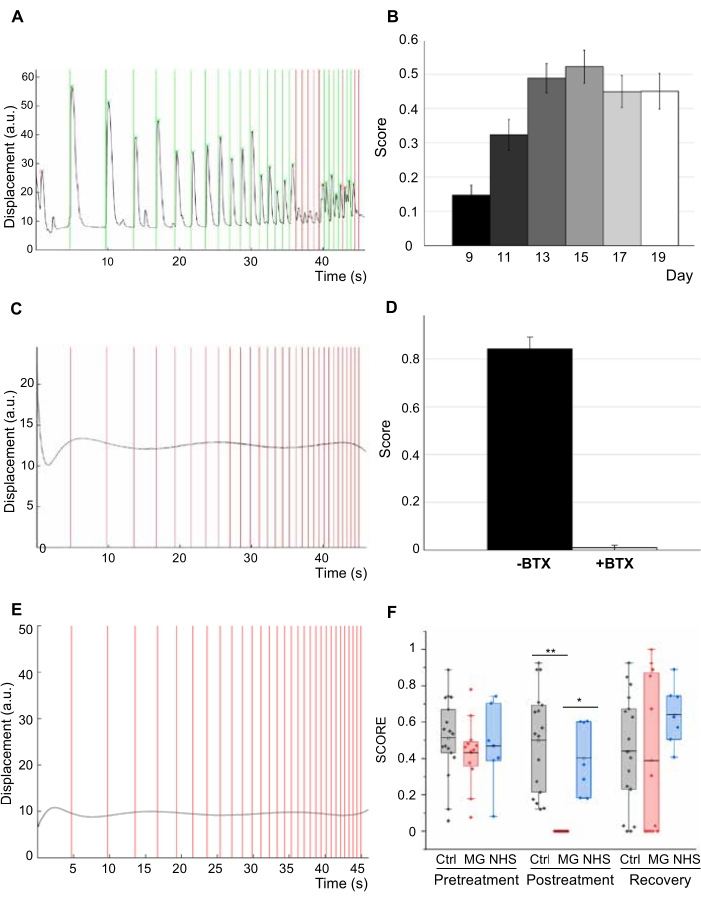
Figure 4. NMJ function. (A) Contractility trace for 3D engineered human NMJs. (B) Score of NMJs in response to light, showing improvement of function over 11 days (n = 47, one-way ANOVA p = 1 x 10-8). Error bars = SEM. (C) Contractility trace after 20 min treatment with 5 mg/mL of the neurotoxin α-bungarotoxin (BTX). (D) Quantification of NMJ function (score) before and after treatment with BTX (n = 6; one-way ANOVA F = 9 x 10-8). (E) Contractility trace of tissues with 20% MG sera after 48 h of treatment. (F) NMJ function assessment for treated and control groups before and after treatment and post-recovery (n = 12; after treatment post-hoc ANOVA F = 0.0002; *indicates p = 0.0015 **indicates p = 3.5 x10−6) (MG = Myasthenia Gravis; NHS=normal human serum). n indicates the number of biological replicates. This figure has been modified with permission from Vila et al.18,19. Please click here to view a larger version of this figure.
Table 1. Myogenic and neural media formulations. Organized list of growth factors and supplements for media. Please click here to download this Table.
Table 2. List of MATLAB functions. Brief explanations of MATLAB functions used for image analysis. This table has been modified with permission from Vila et al.18. Please click here to download this Table.
Supplemental Files
Supplementary Figure 1. Myotube and motoneuron differentiations. (A) Myotube differentiation protocol (MM = maturation media). (B) Immunofluorescence (IF) images showing expression of skeletal muscle markers myosin heavy chain (MHC), α-actinin, and desmin in differentiated myotubes. (C) Motoneuron differentiation protocol. (D) Localization of channelrhodopsin-2 with YFP (ChR2-YFP) to membrane in optogenetic iPSCs and (E) MNs confirming successful infection. (F) Co-expression of choline acetyltransferase (ChAT, red) and ChR2 (green) in optogenetic MNs. Scale bars 100 μm. Reproduced with permission from Vila et al.18. Please click here to download this File.
Supplementary Figure 2. Incorrect processing that requires adjustment of parameters. (A) Incorrect baseline time results in peaks of the wrong shape. (B) A peak threshold that is too high results in peaks being undetected. (C) Too lenient conditions for the detection of minima (i.e., too low values chosen for minMinProminence and/or minMinWidth) result in minima being labeled in the middle or even top of the peaks, therefore being counted as "untriggered" because they start too far from the preceding light pulse. (D) Too stringent conditions for the detection of minima result in the start of the peak being labeled before the light pulse or even missing, as in this example. Reproduced with permission from Vila et al.18. File showing 3D model of bioreactor platform that can be edited and exported to make molds for bioreactor manufacturing using PDMS. Please click here to download this File.
https://github.com/ofvila/NMJ-function-analysis
Supplementary CAD File. File showing 3D model of bioreactor platform that can be edited and exported to make molds for bioreactor manufacturing using PDMS. Please click here to download this File.
