Ribozom profilleme tekniği (RIBO-seq), Kaliforniya Üniversitesi, San Francisco1’dekiJonathan Weissman’ın laboratuvarında geliştirilmiştir. Ribo-seq, gen ekspresyonunu çeviri düzeyinde incelemek için kullanılan diğer yöntemlere kıyasla, mRNA’ya her ribozom bağlamasına odaklanır ve konumu ve transkriptteki ribozomların göreceli sayısı hakkında bilgi sağlar. Protein sentezi sürecinin in vivo olarak izlenmesini sağlar ve hem bireysel mRNA’da hem de hücredeki tüm transkriptom boyunca ribozom yoğunluğunun ölçülmesine izin veren tek kodon çözünürlüğü ve doğruluğu sağlayabilir. RIBO-seq tekniğinin temelinde, çeviri sırasında ribozomların mRNA molekülünü bağlaması ve böylece transkriptin gömülü parçasını bir ribonükleaz sindiriminden koruması yatmaktadır. Ribonokleaz ek olarak, korunmasız mRNA sindirilir ve ribozomlar tarafından çevrelenmiş parçalar – tipik olarak ~ 28-30 nt uzunluğunda – bozulmadan kalır. Ribozomal ayak izleri (RF) olarak adlandırılan bu parçalar daha sonra izole edilebilir, sıralanabilir ve ribozomların tam konumunun tespit edilmesiyle ortaya çıktıkları transkript üzerine eşlenebilir. Aslında, ribozom yeteneği mRNA parçalarını korumak için ribozom yeteneği 1960’lardan beri ribozomal bağlama ve çeviri başlatma alanlarını (Tİs) incelemek içinkullanılmaktadır 2,3,4. Bununla birlikte, derin sıralama teknolojisindeki ilerleme ile RIBO-seq, ribozom katılımı yoluyla protein sentezi 6 hakkında genom çapında bilgi sağlayabilen çeviri izleme5 için altın bir standarthalinegelmiştir. Ribozom profilleme, transkriptom ve proteom6’nınölçülmesi arasında var olan teknolojik boşluğu doldurdu.
Ribozom profilleme yapmak için, araştırılan koşullar altında büyüyen organizmanın hücre lizatını elde etmeliyiz. Hücre toplama ve lizis sırasında bu koşulların bozulması güvenilmez veriler sağlayabilir. Bunu önlemek için, çeviri inhibitörleri, hızlı hasat ve sıvı azot içinde flaş dondurma yaygın olarak kullanılır. Hücreler, mikser değirmeni7,8 veya boncuk çırpıcı9gibi mekanik bir homojenizatörde kriyojenik taşlama ve pipet10 veya iğne ile tritürasyon ile11ile yutabilir. Lizis tamponu, hücrelerin pulverizasyondan hemen önce veya kısa bir süre sonra eklenebilir. Protokolümüzde, harç ve pestle’yi ön soğutmak için sıvı nitrojenin yanı sıra, sonifikasyon gibi yöntemler uygulandığında sıklıkla karşılaşılan RNA makasını önleyen bakteri hücre duvarının bozulmasına daha nazik bir yaklaşım olarak alüminyum oksit kullanıyoruz. Pulverizasyondan sonra harcın soğutulmuş içeriğine buz gibi bir lizis tamponu ekliyoruz. Uygun bir lizis tamponunun seçilmesi ribozomal ayak izlerinin en iyi çözünürlüğünü elde etmek için önemlidir. İyonik mukavemet hem RF boyutunu hem de okuma çerçevesi hassasiyetini etkilediğinden, tampon bileşiminin mRNA’lar11,12’deribozomal doluluğu etkilemediği görünse bile, düşük iyonik mukavemet ve tampon kapasitesine sahip lizis tamponlarının kullanılması önerilir. Lizis tamponunun önemli bileşenleri, varlığı ribozomal alt ünüllerin ayrışmasını önleyen ve bakteriyel ribozomlardaki spontan konformasyonel değişiklikleri inhibe eden magnezyum iyonlarıdır11,13. Kalsiyum iyonları da önemli bir rol oynar ve bakteriyel ribozom profilleme yönteminde kullanılan mikrokoksal çekirdeğin (MNaze) aktivitesi için gereklidir14. Guanosine ilavesi 5′-[β,γ-imido]trifosfat (GMP-PNP), GTP’nin hidrolize edilemez bir analogu, kloramfenikol ile birlikte liziz sırasında çeviriyi engeller15.
Lysate elde edildiğinde, santrifüjleme ile netleştirilir ve her biri bir RIBO-seq ve aynı anda gerçekleştirildikleri için yüksek verimli toplam mRNA dizilimi (RNA-seq) için iki bölüme ayrılır (Şekil 1). RNA-seq, veri analizi sırasında hem RIBO-seq hem de RNA-seq’ten gelen verilerin karşılaştırılmasına olanak sağlayan bir referans noktası sağlar. Araştırılan translatom, ribozomal ayak izlerinin mRNA bolluğuna normalleştirilmesi ile tanımlanır16. RNA-seq’ten gelen veriler,17kopyalama veya sıralama yapıtlarını tanımlamaya da yardımcı olabilir.
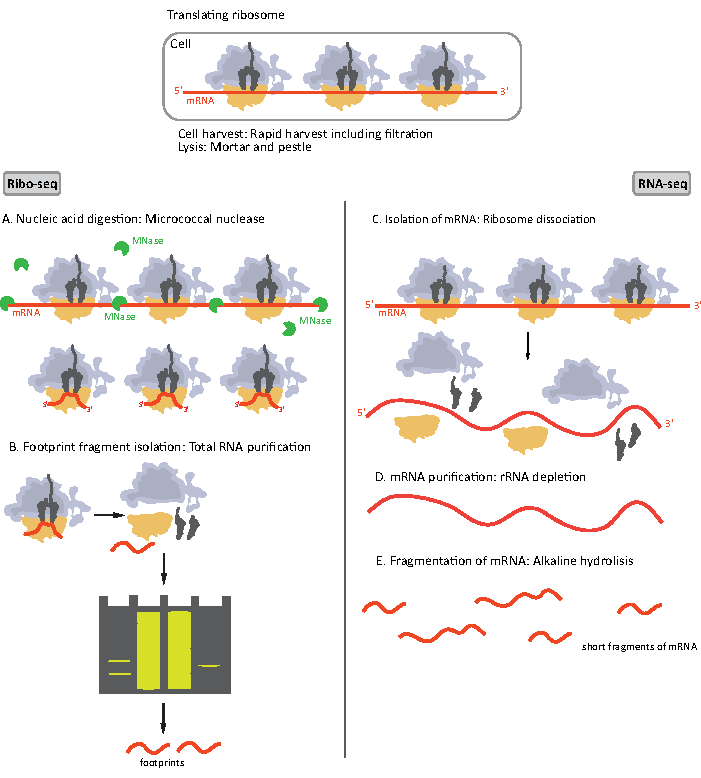
Şekil 1. RIBO-seq ve RNA-seq için mRNA örnek hazırlama şemaları. RIBO-seq kitaplığı hazırlanması için RNA, MNase (A) ile sindirilir ve ardından ~28-30 nt uzunluğunda (B) RF boyut seçimi; RNA-seq RNA yalıtılır (C), rRNA (D) tükenmiş ve elde edilen mRNA rastgele değişen uzunluklarda (E) parçalara bölünmüş. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
RIBO-seq ve RNA-seq için numune hazırlama prosedürünün ilk adımları biraz farklıdır (Şekil 1). Ribozomal profilleme için, ribozomlar tarafından korunmayan mRNA moleküllerini bozmak için lizatın belirli bir endonokleaz tarafından sindirilmesi gerekir. Standart protokollerde, elde edilen monozomlar bir sakkaroz yastığı ultrasantrifüjasyonu veya sakkaroz gradyan ultracentrifugation8,14ile kurtarılır. Bu yazıda, bu adımın bakterilerde RIBO-seq için gerekli RF’yi izole etmek için gerekli olmadığını, aynı şekilde ökaryotik hücreler için18, ve poliakrilamid jelden uygun uzunlukta mRNA parçalarının boyut seçiminin yeterli olduğunu gösteriyoruz.
RNA-seq için mRNA, rRNA’nın toplam RNA’dan tükenmesi ile elde edilir – rRNA molekülleri, streptavidin kaplı manyetik boncuklara bağlanan biyotinillenmiş oligonükleotid problarına melezleşir. RRNA-oligonükleotid-boncuk kompleksleri daha sonra bir mıknatıs ile numuneden çıkarılır ve rRNA tükenmiş bir örnek19,20ile sonuçlanır. Saflaştırılmış mRNA molekülleri daha sonra alkali hidroliz ile rastgele parçalanır. Elde edilen mRNA parçalarının yanı sıra ribozomal ayak izleri cDNA kütüphanelerine dönüştürülür ve derin sıralama için hazırlanır (Şekil 2). Bu, alkali hidroliz (mRNA için) ve enzimatik sindirimden (RF için) sonra ihtiyaç duyulan uç onarımını içerir: 3′ uçların defosforilasyonu ve ardından 5′ uçların fosforilasyonu. Sonraki adımlar adaptör ligasyonu ve Illumina platformunu kullanarak yeni nesil sıralama (NGS) için gereken sıralarla çerçevelenmiş cDNA eklemeleri oluşturmak için ters transkripsiyondur. Kütüphane hazırlığının son aşaması, yapıların bir kanalda çeşitli örneklerin çoklayıcı ve sıralı olmasına izin vermek için örnek özel barkodlarla yükseltildiği ve etiket edildiği bir PCR reaksiyondur. Sıralamadan önce, kütüphanelerin kalitesi ve miktarı çip üzerindeki yüksek hassasiyetli DNA elektroforez ile değerlendirilir. Uygun parametrelere sahip cDNA kitaplıkları daha sonra havuza alınıp sıralanabilir. Sıralama, kitaplık sayısına, gerekli okuma uzunluğuna ve sıralama derinliğine bağlı olarak MiSeq, NextSeq veya HighSeq gibi farklı Illumina platformlarında gerçekleştirilebilir. Sıralamadan sonra biyoinformatik analiz yapılır.

Şekil 2. Kütüphane hazırlığı. Kütüphane hazırlığı, uç onarımını, adaptör ligasyonunu, ters transkripsiyonu ve barkodlama ile amplifikasyonu içerir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Ribozom profilleme, bilimsel soruya göre kolayca değiştirilebilen ve ayarlanabilen evrensel bir yöntemdir. Başlangıçta maya1’dekullanıldı, ancak bakteri hücrelerine uygulandıktan kısa bir süre sonra21 ve fare10, zebra balığı22,meyve sineği 23 ve Arabidopsis thaliana24dahil olmak üzere ökaryotik model organizmalara uygulandı. Ayrıca farklı ribozom türlerini incelemek için de kullanılmıştır: sitoplazmik, mitokondriyal25,26 ve kloroplast27,28. Ökaryotlarda RIBO-seq, başlatma 10 , 11 , 29 , 30 ,31,32,uzama1,10 , 11,31,33,ribozom oyalama 33 ve uygunlukdeğişikliği33 dahil olmak üzere çevirinin belirli yönlerini araştırmak için yaygın olarak uyarlanmış ve rafine edilmiştir. Değişikliklerin çoğu farklı çeviri inhibitörlerinin kullanımını içerir. Bununla birlikte, bakterilerde, gerekli etki mekanizmasına sahip inhibitörlerin azlığı nedeniyle benzer çalışmaların yapılması zor olmuştur34. Bakterilerde en sık kullanılan çeviri inhibitörü, peptidyl transferaz merkezine (PTC) bağlanan ve A bölgesinde aminoasil-tRNA’nın doğru konumlandırılmasını önleyen kloramfenikoldür (CAM). Sonuç olarak, CAM, uzun ribozomların tutuklanmasına yol açan bir peptit bağı oluşumunu önler35. Bakterilerdeki diğer çeviri inhibitörleri tetrasiklin (TET)36, retapamulin (RET)34 ve Onc11237’dir. A-sitedeki tRNA’nın antikon kök döngüsü ile doğrudan örtüşerek ribozom için tRNA doğumunu önleyen TET, başlangıçta CAM tedavisinden elde edilen sonuçları doğrulamak için uygulanmıştır, çünkü her ikisi de çeviri uzamasını engelleyen antibiyotiklerdir38. TET’nin birincil Tİs’i tespit etmesi bulundu, ancak dahili Tİs36’yıortaya koyamadı. RET, bakteriyel ribozom PTC’sinde bağlanır ve A bölgesinde bir uzay aminoasil-tRNA ile müdahale ederek ilk peptit bağının oluşumunu önler. RET uygulamak, hem birincil hem de dahili TISs34’teribozomların tutuklanmasıyla sonuçlanır. Prolin bakımından zengin bir antimikrobiyal peptit olan Onc112, çıkış tüneline bağlanır ve ribozomal A bölgesinde aminoasil-tRNA bağlamasını engeller. Sonuç olarak, Onc112 başlatma komplekslerinin uzama aşaması37’yegirmesini önler.
Ribozom profillemenin sağladığı ana bilgi ribozom yoğunluğu ve mRNA üzerindeki konumlarıdır. Çeşitli büyüme koşullarında çeviri düzeyinde diferansiyel gen ekspresyonunun araştırılması 1,6 , çeviri verimliliğininölçülması 1,38,39 ve ribozomal duraklama10gibi çeviri düzenleme olaylarını tespit etmek için başarıyla uygulanmıştır. RIBO-seq ayrıca, yeni ve / veya çok kısa protein kodlama genlerinin tanımlanmasına yol açan açıklamalı ncRNA, psödojenler ve açıklamalanmamış küçük açık okuma çerçevelerinin (ORF) çevirisini ortaya çıkarmaya izin verir10,12,22,30,37. Bu gibi durumlarda RIBO-seq genom ek açıklamalarını ince ayar yapabilir ve iyileştirebilir. Çevrilmiş ORF’lerin tanımlanmasına ve nicel doğasına yönelik yüksek hassasiyeti ile ribozom profilleme, proteom tayini için veya proteomik çalışmalara yardımcı olmak için bir vekil görevi de edebilir31,34,39. Ribozom profilleme, Tİs’i haritalayarak bilinen proteinlerin N-terminal genişletilmiş ve kesilmiş izoformlarını ortaya koymaktadır10,32. RIBO-seq ayrıca14 , 21,24proteinlerinin ortak çevirisel katlamasını incelemek için uyarlanmıştır. Buyöntem,1 ,10,39 uzama hızlarının veya bireysel kodonların kod çözme hızlarının ölçülmesine olanak tanır 6 veçevirinin nicel modellerinin geliştirilmesine yardımcı olur17. Ribozom profilleme yöntemi ayrıca7, 15,17,frameshifting 40,stop-codon readthrough 21 , sonlandırma/geri dönüşüm kusurları 41,42ve ribozomal konformasyon değişiklikleri33 ökaryotlarda ribozom duraklaması hakkında mekanistik içgörüler sağlayabilir. RIBO-seq ayrıca 16,43ökaryotlarda miRNAs6 ve RNA bağlayıcı proteinler gibi spesifik trans-etkili faktörlerin çeviri üzerindeki etkisini incelemek için uyarlanmıştır. Bununla birlikte, deneysel tasarımın ve RIBO-seq’in elde edilen çözünürlüğünün, elde edilen sıralama verilerinden çıkarabilecek bilgi miktarını belirlediğini kabul etmek önemlidir12.
