טכניקת פרופיל הריבוזום (RIBO-seq) פותחה במעבדתו של ג’ונתן וייסמן באוניברסיטת קליפורניה בסן פרנסיסקו1. בהשוואה לשיטות אחרות המשמשות לחקר ביטוי גנים ברמה התרגומית, RIBO-seq מתמקד בכל מחייב ריבוזום ל- mRNA ומספק מידע על מיקומו ועל המספר היחסי של ריבוזומים בתעתיק. זה מאפשר ניטור התהליך של סינתזת חלבון vivo והוא יכול לספק רזולוציה קודון יחיד ודיוק המאפשר מדידה של צפיפות ריבוזום על שניהם, mRNA הפרט לאורך התמלול כולו בתא. בבסיס טכניקת RIBO-seq טמונה העובדה כי במהלך התרגום הריבוזום קושר את מולקולת mRNA ובכך מגן על החלק הקבור של התמליל מעיכול ריבונוקלאאז. עם תוספת של ribonuclease, mRNA לא מוגן מתעכל ואת שברים מוקף ריבוזומים – בדרך כלל של ~ 28-30 nt ארוך – להישאר שלם. שברים אלה, הנקראים עקבות ריבוזומליות (RF), יכולים להיות מבודדים, רציפים וממופים לתעתיק שמקורו בזיהוי המיקום המדויק של הריבוזומים. למעשה, היכולת הריבוזום להגן על שברי mRNA שימש מאז 1960 ללמוד אתרי ייזום מחייב ותרגום ריבוזומלי (TIS)2,3,4. עם זאת, עם ההתקדמות בטכנולוגיית רצף עמוק, RIBO-seq הפך תקן זהב לניטור תרגום5 אשר, באמצעות מעורבות ריבוזום, יכול לספק מידע גנומי רחב על סינתזת חלבונים6. פרופיל ריבוזום מילא את הפער הטכנולוגי שהיה קיים בין כימות התמלול לבין פרוטאום6.
כדי לנהל פרופיל ריבוזום אנחנו צריכים להשיג ליזאט התא של האורגניזם שגדל בתנאים שנחקרו. שיבוש תנאים אלה במהלך איסוף תאים ותמוגה עשוי לספק נתונים לא אמינים. כדי למנוע זאת, מעכבי תרגום, קציר מהיר והקפאת פלאש בחנקן נוזלי משמשים בדרך כלל. תאים יכולים להיות lysed על ידי שחיקה קריוגנית homogenizer מכני כמו טחנת מערבל7,8 או מקצף חרוזים9, ועל ידי טריטורציה באמצעות פיפטה10 או עם מחט11. מאגר התמוגה ניתן להוסיף רק לפני או זמן קצר לאחר ריסוק של התאים. בפרוטוקול שלנו אנו משתמשים חנקן נוזלי כדי מרגמה precool ועלי, כמו גם תחמוצת אלומיניום כגישה עדינה יותר להפרעה של דופן התא החיידקי, אשר מונע גזירה RNA לעתים קרובות נתקל כאשר שיטות כגון sonification מוחלים. לאחר ריסוק, אנו מוסיפים חיץ תמוגה קר כקרח לתכולת המרגמה מקוררת. בחירה של מאגר תמוגה מתאים חשוב להשגת הרזולוציה הטובה ביותר של עקבות ריבוזומליות. מאז חוזק יוני משפיע הן על גודל RF ואת דיוק מסגרת הקריאה, מומלץ כיום להשתמש מאגרי תמוגה עם חוזק יוני נמוך קיבולת חיץ, גם אם נראה כי הרכב המאגר אינו משפיע על תפוסה ריבוזואלית על mRNAs11,12. מרכיבים חשובים של מאגר התמוגה הם יוני מגנזיום, שנוכחותם מונעת ניתוק של יחידות המשנה הריבוזומליות ומעכבת שינויים קונפורמיים ספונטניים בריבוזומים החיידקיים11,13. יוני סידן גם לשחק תפקיד משמעותי והם חיוניים לפעילות של גרעין מיקרוקוקאלי (MNase) המשמש בשיטת פרופיל ריבוזום חיידקי14. תוספת של גואנוסין 5′-[β,γ-imido]טריפוספט (GMP-PNP), אנלוגי שאינו הידרוליזה של GTP, יחד עם כלוראמפניקול מעכב תרגום במהלך תמוגה15.
כאשר מתקבל הליסנט, מובהר על ידי צנטריפוגה ומחולק לשני חלקים, כל אחד עבור RIBO-seq ורצף mRNA כולל בעל תפוקה גבוהה (RNA-seq) שכן הם מבוצעים בו זמנית (איור 1). RNA-seq מספק נקודת התייחסות המאפשרת השוואה של נתונים הן RIBO-seq והן RNA-seq במהלך ניתוח נתונים. התרגום שנחקר מוגדר על ידי נורמליזציה של עקבות ריבוזומליות לשפע mRNA16. נתונים מ-RNA-seq יכולים גם לסייע בזיהוי שיבוט או רצף של חפצים17.
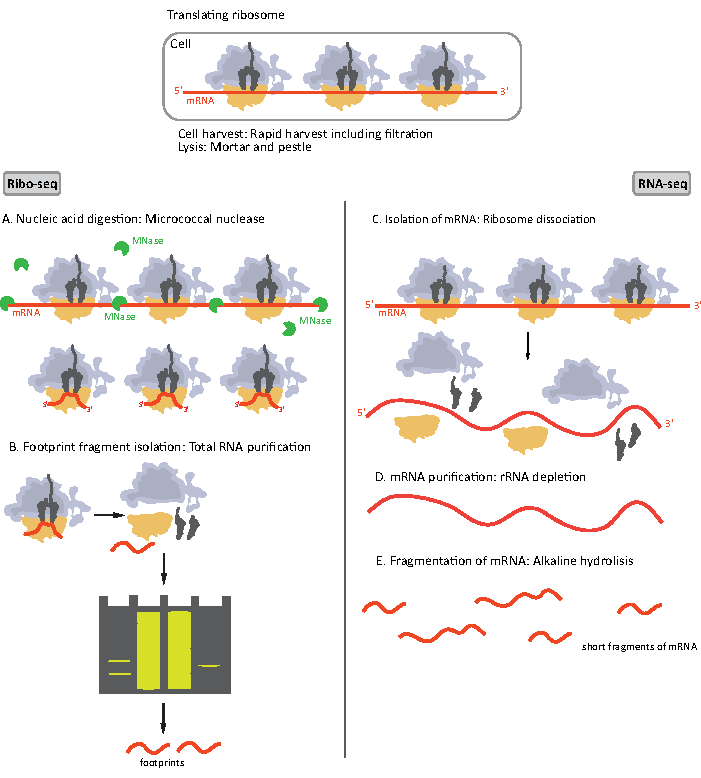
איור 1. שרטוטים של הכנת דגימת mRNA עבור ריבו-seq ו- RNA-seq. להכנת ספריית RIBO-seq, RNA מתעכל עם MNase (A), ואחריו מבחר הגודל של RF של ~ 28-30 nt אורך (B); עבור RNA-seq RNA מבודד (C), מדולדל של rRNA (D), ואת mRNA וכתוצאה מכך הוא מקוטע באופן אקראי לרסיסים באורכים משתנים (E). לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
השלבים הראשוניים של הליך ההכנה לדוגמה עבור RIBO-seq ו- RNA-seq שונים במקצת (איור 1). עבור פרופיל ריבוזומלי, lysate צריך להיות מתעכל על ידי אנדונוקלאאז מסוים כדי להשפיל את מולקולות mRNA לא מוגן על ידי ריבוזומים. בפרוטוקולים סטנדרטיים, מונוזומים המתקבלים הם התאוששו על ידי אולטרה צנטריפוגציה כרית סוכרוז או אולטרה צנטריפוגציה שיפוע סוכרוז8,14. במאמר זה, אנו מראים כי שלב זה אינו הכרחי כדי לבודד RF הנדרש עבור RIBO-seq בחיידקים, כמו כן עבור תאים אוקריוטיים18, וכי מבחר גודל של שברי mRNA אורך המתאים מן ג’ל polyacrylamide מספיק.
עבור RNA-seq, mRNA מתקבל על ידי דלדול של rRNA מן RNA הכולל – מולקולות rRNA הכלאה לבדיקות אוליגונוקלאוטיד biotinylated אשר נקשרים חרוזים מגנטיים מצופים סטרפטבידין. קומפלקסים rRNA-oligonucleotide-חרוזים מוסרים לאחר מכן מן המדגם עם מגנט וכתוצאה מכך מדגם מדולדל rRNA19,20. מולקולות mRNA מטוהרים לאחר מכן מפוצלים באופן אקראי על ידי הידרוליזה אלקליין. שברי ה-mRNA המתקבלים, כמו גם עקבות הריבוזומליות, מומרים לספריות cDNA ומוכנים לריצוף עמוק(איור 2). זה כרוך בתיקון מסתיים הדרוש לאחר הידרוליזה אלקליין (עבור mRNA) ועיכול אנזימטי (עבור RF): dephosphorylation של 3 ‘ קצוות ואחריו זרחון של קצוות 5 ‘. השלבים הבאים הם קשירת מתאמים ואת התמלול ההפוך כדי ליצור מוסיף cDNA ממוסגר על ידי רצפים הנדרשים עבור ריצוף הדור הבא (NGS) באמצעות פלטפורמת Illumina. השלב האחרון של הכנת הספרייה הוא תגובת PCR שבה המבנים מוגברים ומתויגים עם ברקודים ספציפיים לדוגמה כדי לאפשר multiplexing רצף דגימות שונות בערוץ אחד. לפני הרצף, האיכות והכמות של הספריות מוערכים על ידי אלקטרופורזה DNA רגישות גבוהה על השבב. לאחר מכן ניתן לאגד ולרצף ספריות cDNA עם פרמטרים מתאימים. ניתן לבצע רצף בפלטפורמות שונות של Illumina, כגון MiSeq, NextSeq או HighSeq, בהתאם למספר הספריות, אורך הקריאה הנדרש ועומק הרצף. לאחר הרצף מבוצע הניתוח הביואינפורמטי.

איור 2. הכנת ספרייה. הכנת הספרייה כוללת את תיקון הקצוות, קשירת מתאמים, שעתוק הפוך והגברה עם ברקודינג. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
פרופיל הריבוזום הוא שיטה אוניברסלית אשר ניתן לשנות ולהתאים בקלות על פי השאלה המדעית. במקור הוא שימש שמרים1, אבל זמן קצר לאחר מכן הוחל על תאים חיידקיים21, כמו גם אורגניזמים מודל אוקריוטי כולל עכבר10, זברה דג22, זבוב פירות23 ו Arabidopsis thaliana24. הוא שימש גם לחקר סוגים שונים של ריבוזומים: ציטופלסמי, מיטוכונדריאלי25,26 וכלורופלסט27,28. באיקריוטים RIBO-seq מותאם ומעודן בדרך כלל כדי לחקור היבטים ספציפיים של התרגום, כולל חניכה10,11,29,30,31,32, התארכות 1,10,11,31,33, ריבוזום השתהות33 ושינוי קונפורמציה33. רוב השינויים כרוכים בשימוש במעכבי תרגום שונים. בחיידקים עם זאת, מחקרים מקבילים היו קשים לביצוע בגלל מחסור במעכבים עם מנגנון הפעולה הנדרש34. מעכב התרגום הנפוץ ביותר בחיידקים הוא כלוראמפניקול (CAM) הנקשר למרכז הפפטידיל טרנספראז (PTC) ומונע מיקום נכון של האמינואציל-tRNA באתר A. כתוצאה מכך, CAM מונע היווצרות של קשר פפטיד אשר מוביל למעצר ריבוזומים מוארכים35. דוגמאות אחרות של מעכבי תרגום בחיידקים הם טטרציקלין (TET)36, retapamulin (RET)34 ו Onc11237 אשר שימשו כדי לחקור אתרי ייזום תרגום. TET, אשר מונע משלוח tRNA לריבוזום על ידי חפיפה ישירה עם לולאת גזע אנטיקודון של tRNA באתר A, הוחל במקור כדי לאמת את התוצאות שהתקבלו מטיפול CAM שכן שניהם אנטיביוטיקה מעכבת התארכות תרגום38. TET נמצא לזהות TIS ראשי, עם זאת לא הצליח לחשוף פנימי TIS36. RET נקשר PTC של ריבוזום חיידקי, ומונע היווצרות של הקשר פפטיד הראשון על ידי הפרעה עם aminoacyl-tRNA מוארך באתר. החלת תוצאות RET במעצר ריבוזומים הן ראשי, כמו גם פנימי TISS34. Onc112, פפטיד אנטי מיקרוביאלי עשיר בפרולין, נקשר במנהרת היציאה וחוסם את כריכת aminoacyl-tRNA באתר ריבוזומלי A. כתוצאה מכך, Onc112 מונע ממתחמי חניכה להיכנס לשלב התארכות37.
המידע העיקרי פרופיל ריבוזום מספק הוא צפיפות ריבוזומים ואת מיקומם על mRNA. זה יושם בהצלחה לחקור ביטוי גנים דיפרנציאליים ברמת התרגום בתנאי צמיחה שונים1,6, למדוד יעילות תרגום1,38,39 ולזהות אירועי ויסות תרגום כגון השהייה ריבוזומלית10. RIBO-seq מאפשר גם לחשוף את התרגום של ncRNA ביאורים, פסאודוגנים ומסגרות קריאה פתוחות קטנות ללא ביאורים (ORF) המוביל לזיהוי של גנים קידוד חלבון הרומן ו / או קצר מאוד10,12,22,30,37. במקרים כאלה, RIBO-seq יכול לכוונן ולשפר ביאור הגנום. עם הרגישות הגבוהה שלה לזיהוי של ORFs מתורגם ואופיו הכמותי, פרופיל ריבוזום יכול לשמש גם פרוקסי לקביעת פרוטאום או בסיוע מחקרים פרוטאומיקים31,34,39. על ידי מיפוי TIS, פרופיל ריבוזום חושף N-סופני מורחב וקצץ isoforms של חלבונים ידועים10,32. RIBO-seq הותאם גם ללמוד קיפול תרגום משותף של חלבונים14,21,24. שיטה זו מאפשרת מדידה של שיעורי התארכות1,10,39 או מהירויות פענוח של קודונים בודדים6 ומסייעת בפיתוח מודלים כמותיים של תרגום17. שיטת פרופיל ריבוזום הוא גם מסוגל לספק תובנות מכניות לתוך הריבוזום השהיה בחיידקים7,15,17, frameshifting40, עצור קודון readthrough21, סיום / מיחזור פגמים41,42 ו שינויים קונפורמציה ריבוזומלית33 באיקריוטים. RIBO-seq הותאם גם כדי לבחון את ההשפעה של גורמים טרנס-משחקים ספציפיים על תרגום כגון miRNAs6 וחלבונים מחייבי RNA באיקריוטים16,43. עם זאת, חשוב להכיר בכך שהתכנון הניסיוני והרזולוציה המתקבלת של RIBO-seq קובעים את כמות המידע שניתן לחלץ מנתוני הרצףהמתקבלים 12.
