Ribosomprofileringsteknikken (RIBO-seq) ble utviklet i laboratoriet til Jonathan Weissman ved University of California, San Francisco1. Sammenlignet med andre metoder som brukes til å studere genuttrykk på translasjonsnivå, fokuserer RIBO-seq på hvert ribosome som binder seg til mRNA og gir informasjon om plasseringen og det relative antallet ribosomer på en transkripsjon. Det muliggjør overvåking av prosessen med proteinsyntese in vivo og kan gi enkel kodonoppløsning og nøyaktighet som tillater måling av ribosomets tetthet på begge, den enkelte mRNA og langs hele transkripsjonen i cellen. Ved grunnlaget for RIBO-seq-teknikken ligger det faktum at ribbeosomet under oversettelse binder mRNA-molekylet og dermed beskytter det begravde fragmentet av transkripsjonen fra en ribonuklease fordøyelse. Ved tilsetning av ribonuklease fordøyes den ubeskyttede mRNA og fragmentene vedlagt av ribosomer – vanligvis ~ 28-30 nt lange – forblir intakte. Disse fragmentene, kalt ribosomale fotavtrykk (RF), kan deretter isoleres, sekvenseres og kartlegges på transkripsjonen de stammer fra, noe som resulterer i deteksjon av ribosomenes nøyaktige posisjon. Faktisk har ribososom evnen til å beskytte mRNA fragmenter blitt brukt siden 1960-tallet for å studere ribosomal binding og oversettelse initieringssteder (TIS)2,3,4. Men med fremskrittet i dyp sekvenseringsteknologi har RIBO-seq blitt en gullstandard for oversettelsesovervåking5 som gjennom ribosomets engasjement kan gi en genom-bred informasjon om proteinsyntese6. Ribosomprofilering fylte det teknologiske gapet som eksisterte mellom kvantifisering av transkripsjonen og proteomet6.
For å gjennomføre ribosomprofilering må vi få cellelys av organismen som hadde vokst under de undersøkte forholdene. Å forstyrre disse forholdene under celleinnsamling og lysis kan gi upålitelige data. For å forhindre dette brukes ofte oversettelseshemmere, rask høsting og blitsfrysing i flytende nitrogen. Celler kan lyses ved kryogen sliping i en mekanisk homogenisator som en miksermølle7,8 eller en perle beater9, og ved triturasjon gjennom en pipette10 eller med en nål11. Lysisbufferen kan tilsetts like før eller kort tid etter pulverisering av cellene. I vår protokoll bruker vi flytende nitrogen til prekolmørtel og pestle, samt aluminiumoksid som en mildere tilnærming til forstyrrelse av bakteriell cellevegg, noe som forhindrer RNA-skjæring ofte oppstår når metoder som sonifisering påføres. Etter pulverisering legger vi til en iskald lysisbuffer i det avkjølte innholdet i mørtelet. Valg av en passende lysisbuffer er viktig for å oppnå den beste oppløsningen av ribosomale fotavtrykk. Siden ionstyrke påvirker både RF-størrelsen og leserammepresisjonen, anbefales det for tiden å bruke lysisbuffere med lav ionisk styrke og bufferkapasitet, selv om det ser ut til at buffersammensetningen ikke påvirker ribosomal belegg på mRNAs11,12. Viktige komponenter i lysisbufferen er magnesiumioner, hvis tilstedeværelse forhindrer dissosiasjon av ribosomale underenheter og hemmer spontane konformasjonsendringer i bakterielle ribosomer11,13. Kalsiumioner spiller også en betydelig rolle og er avgjørende for aktiviteten til mikrokokkkjerne (MNase) som brukes i bakteriell ribosomprofileringsmetode14. Tillegg av guanosin 5′-[β,γ-imido]triphosfat (GMP-PNP), en ikke-hydrolysert analog av GTP, sammen med kloramfenikol hemmer oversettelse under lysis15.
Når lysatet oppnås, avklares det ved sentrifugering og deles inn i to deler, hver for en RIBO-seq og en total mRNA-sekvensering med høy gjennomstrømning (RNA-seq) siden de utføres samtidig (figur 1). RNA-seq gir et referansepunkt som gjør det mulig å sammenligne data fra både RIBO-seq og RNA-seq under dataanalyse. Den undersøkte translatome er definert ved normalisering av ribosomale fotavtrykk til mRNA overflod16. Data fra RNA-seq kan også bidra til å identifisere kloning eller sekvensering av artefakter17.
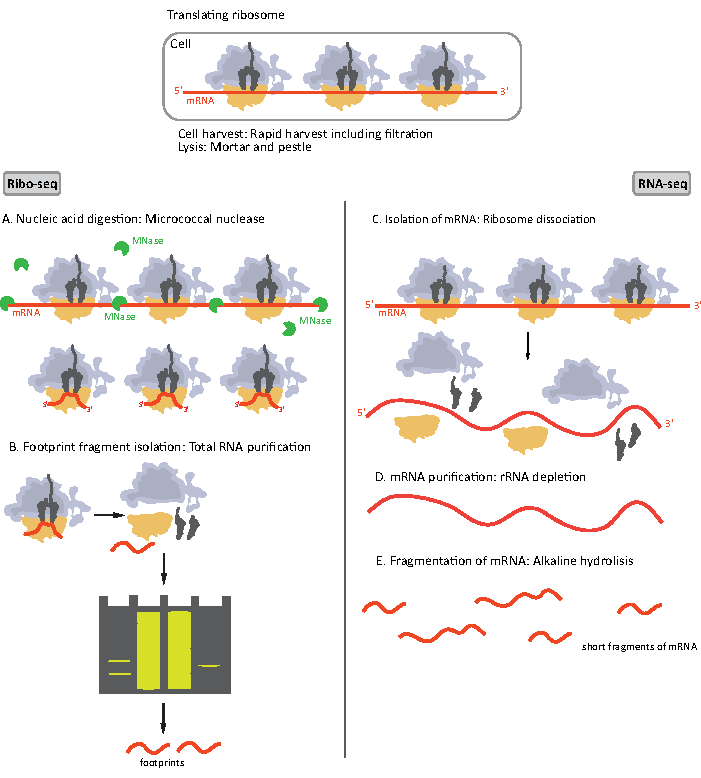
Figur 1. Skjema for mRNA prøvepreparering for RIBO-seq og RNA-seq. For RIBO-seq bibliotek forberedelse fordøyes RNA med MNase (A), etterfulgt av størrelsesvalget av RF på ~ 28-30 nt lengde (B); for RNA-seq RNA er isolert (C), utarmet av rRNA (D), og den resulterende mRNA fragmenteres tilfeldig i fragmenter av varierende lengde (E). Klikk her for å se en større versjon av denne figuren.
De første trinnene i prosedyren for prøvepreparering for RIBO-seq og RNA-seq varierer noe (figur 1). For ribosomal profilering må lysatet fordøyes av en bestemt endonuklease for å forringe mRNA-molekylene som ikke er beskyttet av ribosomene. I standardprotokoller gjenvinnes de oppnådde monosomene av en sukrosepute ultracentrifugation eller en sukrose gradient ultracentrifugation8,14. I denne artikkelen viser vi at dette trinnet ikke er nødvendig for å isolere RF som kreves for RIBO-seq i bakterier, på samme måte for eukaryote celler18, og at størrelsesvalg av riktig lengde mRNA-fragmenter fra polyakrylamidgelen er tilstrekkelig.
For RNA-seq oppnås mRNA ved uttømming av rRNA fra de totale RNA – rRNA-molekylene hybridiserer til de biotinylerte oligonukleotidsondene som binder seg til de streptavidinbelagte magnetiske perlene. RRNA-oligonukleotid-perlekompleksene fjernes deretter fra prøven med en magnet som resulterer i en rRNA utarmet prøve19,20. De rensede mRNA-molekylene fragmenteres deretter tilfeldig av alkalisk hydrolyse. De oppnådde fragmentene av mRNA samt ribosomale fotavtrykk konverteres til cDNA-biblioteker og fremstilles for dyp sekvensering (figur 2). Dette innebærer reparasjon av ender som trengs etter alkalisk hydrolyse (for mRNA) og enzymatisk fordøyelse (for RF): defosforylering av 3′ ender etterfulgt av fosforylering av 5′ ender. De neste trinnene er adaptere ligation og omvendt transkripsjon for å lage cDNA innsatser innrammet av sekvenser som kreves for neste generasjon sekvensering (NGS) ved hjelp av Illumina plattform. Den siste fasen av bibliotekforberedelse er en PCR-reaksjon der konstruksjonene forsterkes og merkes med prøvespesifikke strekkoder for å tillate multipleksing og sekvensering av ulike prøver på en kanal. Før sekvensering vurderes kvaliteten og mengden av bibliotekene av høysensitiv DNA on-chip elektroforese. cDNA-biblioteker med riktige parametere kan deretter grupperes og sekvenseres. Sekvensering kan utføres på forskjellige Illumina-plattformer, for eksempel MiSeq, NextSeq eller HighSeq, avhengig av antall biblioteker, nødvendig leselengde og sekvenseringsdybde. Etter sekvensering utføres den bioinformatiske analysen.

Figur 2. Klargjøring av biblioteket. Biblioteksforberedelse inkluderer endereparasjon, adaptere ligation, omvendt transkripsjon og forsterkning med barkoding. Klikk her for å se en større versjon av denne figuren.
Ribosomet profilering er en universell metode som lett kan modifiseres og justeres i henhold til det vitenskapelige spørsmålet. Opprinnelig ble den brukt i gjær1, men kort tid etter at den ble brukt på bakterieceller21 samt eukaryote modellorganismer, inkludert mus10, sebrafisk22, fruktflue23 og Arabidopsis thaliana24. Den ble også brukt til å studere forskjellige ribosomtyper: cytoplasmatisk, mitokondrie25,26 og kloroplast27,28. I eukaryoter er RIBO-seq vanligvis tilpasset og raffinert for å undersøke spesifikke aspekter ved oversettelse, inkludert initiering10,11,29,30,31,32, forlengelse1,10,11,31,33, ribosom stalling33 og konformasjonsendring33. De fleste modifikasjonene innebærer bruk av forskjellige oversettelseshemmere. I bakterier har imidlertid analoge studier vært vanskelige å gjennomføre på grunn av hemmeres paucity med den nødvendige virkningsmekanismen34. Den mest brukte oversettelseshemmeren i bakterier er kloramfenikol (CAM) som binder seg til peptidyloverføringssenteret (PTC) og forhindrer riktig posisjonering av aminoacyl-tRNA på A-området. Som et resultat forhindrer CAM dannelsen av en peptidbinding som fører til å arrestere de langstrakte ribosomene35. Andre eksempler på oversettelseshemmere i bakterier er tetracyklin (TET)36, retapamulin (RET)34 og Onc11237 som har blitt brukt til å undersøke oversettelsesinitieringssteder. TET, som forhindrer tRNA-levering til ribosomet ved å overlappe direkte med antikodonstammeløkken til tRNA på A-stedet, ble opprinnelig brukt for å verifisere resultatene oppnådd fra CAM-behandling siden de begge er antibiotika som hemmer oversettelsesforlengelse38. TET ble funnet for å oppdage primær TIS, men kunne ikke avsløre intern TIS36. RET binder seg i PTC av bakteriell ribosom, og forhindrer dannelse av den første peptidbindingen ved å forstyrre en langstrakt aminoacyl-tRNA på A-området. Bruk av RET resulterer i ribosomer arrestasjon på både primære og interne TISs34. Onc112, et prolinrikt antimikrobielt peptid, binder seg i utgangstunnelen og blokkerer aminoacyl-tRNA-binding i ribosomal A-området. Som et resultat forhindrer Onc112 initieringskomplekser fra å komme inn i forlengelsesfasen37.
Hovedinformasjonen ribosomeprofilering gir er ribosomer tetthet og deres posisjon på mRNA. Det ble brukt til å undersøke differensialgenuttrykk på oversettelsesnivå under ulike vekstforhold1,6, måle translasjonell effektivitet1,38,39 og oppdage oversettelsesreguleringshendelser som ribosomal pause10. RIBO-seq gjør det også mulig å avdekke oversettelsen av kommenterte ncRNA, pseudogenes og unannotated små åpne leserammer (ORF) som fører til identifisering av nye og / eller svært korte proteinkodingsgener10,12,22,30,37. I slike tilfeller kan RIBO-seq finjustere og forbedre genommerknaden. Med sin høye følsomhet for identifisering av oversatte ORFer og dens kvantitative natur, kan ribosomprofilering også fungere som proxy for proteombestemmelsen eller i å hjelpe proteomikkstudier31,34,39. Ved å kartlegge TIS avslører ribosomprofilering N-terminalt utvidede og avkortede isoformer av kjente proteiner10,32. RIBO-seq ble også tilpasset for å studere samtidig translasjonell folding av proteiner14,21,24. Denne metoden gjør det mulig å måle forlengelseshastigheter1,10,39 eller dekodingshastigheter for individuelle codons6 og hjelper til med å utvikle kvantitative modeller av oversettelse17. Ribosomet profilering metoden er også i stand til å gi mekanistisk innsikt i ribosomet pause i bakterier7,15,17, frameshifting40, stop-codon readthrough21, terminering / resirkulering defekter41,42 og ribosomal konformasjon endringer33 i eukaryoter. RIBO-seq ble også tilpasset for å undersøke effekten av spesifikke transvirkende faktorer på oversettelse som miRNAs6 og RNA-bindende proteiner i eukaryoter16,43. Det er imidlertid viktig å erkjenne at den eksperimentelle utformingen og den oppnådde oppløsningen av RIBO-seq bestemmer mengden informasjon som kan trekkes ut fra de resulterende sekvenseringsdataene12.
