A técnica de perfil ribossomo (RIBO-seq) foi desenvolvida no laboratório de Jonathan Weissman na Universidade da Califórnia, São Francisco1. Em comparação com outros métodos usados para estudar a expressão genética no nível translacional, RIBO-seq concentra-se em cada ligação ribossosome ao mRNA e fornece informações sobre sua localização e o número relativo de ribossomos em uma transcrição. Permite monitorar o processo de síntese proteica in vivo e pode fornecer resolução e precisão de códon único permitindo a medição da densidade ribossomo em ambos, o mRNA individual e ao longo de todo o transcriptome na célula. Na base da técnica RIBO-seq está o fato de que durante a tradução o ribossomo liga a molécula de mRNA e, assim, protege o fragmento enterrado da transcrição de uma digestão ribonuclease. Após a adição da ribonuclease, o mRNA desprotegido é digerido e os fragmentos fechados por ribossomos – tipicamente de ~28-30 nt de comprimento – permanecem intactos. Esses fragmentos, chamados pegadas ribossômicas (RF), podem então ser isolados, sequenciados e mapeados na transcrição de que se originaram resultando na detecção da posição exata dos ribossomos. De fato, a capacidade ribossômica de proteger fragmentos de mRNA tem sido usada desde a década de 1960 para estudar os locais de iniciação de ligação ribossômica e tradução (TIS)2,3,4. No entanto, com o avanço da tecnologia de sequenciamento profundo, o RIBO-seq tornou-se um padrão ouro para o monitoramento de tradução5 que, através do engajamento ribossomo, pode fornecer uma informação em todo o genoma sobre asíntese deproteínas 6 . O perfil ribossomo preencheu a lacuna tecnológica existente entre quantificar o transcriptome e o proteome6.
Para realizar o perfil ribossomo precisamos obter lisecelulares do organismo que cresceu sob as condições investigadas. Interromper essas condições durante a coleta e lise celular pode fornecer dados não confiáveis. Para evitar isso, inibidores de tradução, colheita rápida e congelamento de flash em nitrogênio líquido são comumente usados. As células podem ser lístidas pela moagem criogênica em um homogeneizador mecânico como um moinho de batedeira7,8 ou um batedor de contas9, e por trituração através de uma pipeta10 ou com uma agulha11. O tampão de lise pode ser adicionado pouco antes ou pouco depois da pulverização das células. Em nosso protocolo usamos nitrogênio líquido para argamassa pré-colono e pilão, bem como óxido de alumínio como uma abordagem mais suave para a interrupção da parede celular bacteriana, o que impede a tesoura de RNA frequentemente encontrada quando métodos como a sonificação são aplicados. Após a pulverização, adicionamos um tampão de lise gelada no conteúdo resfriado da argamassa. A seleção de um tampão de lise apropriado é importante para obter a melhor resolução de pegadas ribossômicas. Uma vez que a força iônica afeta tanto o tamanho do RF quanto a precisão do quadro de leitura, recomenda-se atualmente o uso de tampões de lise com baixa resistência iônica e capacidade de tampão, mesmo que pareça que a composição do buffer não afeta a ocupação ribossômica em mRNAs11,12. Componentes importantes do tampão de lise são íons de magnésio, a presença que impede a dissociação das subunidades ribossômicas e inibe alterações conformais espontâneas nos ribossomos bacterianos11,13. Os íons de cálcio também desempenham um papel significativo e são essenciais para a atividade da nuclease microcócica (MNase) utilizada no método de perfil ribossomo bacteriano14. Adição de guanosina 5′-[β,γ-imido]triphosfato (GMP-PNP), um análogo não hidroglizável do GTP, juntamente com o clorofenicol inibe a tradução durante a lise15.
Quando o lisato é obtido, ele é esclarecido por centrifugação e dividido em duas porções, cada uma para um RIBO-seq e um sequenciamento de mRNA total de alta produtividade (RNA-seq) uma vez que são realizados simultaneamente(Figura 1). O RNA-seq fornece um ponto de referência que permite a comparação de dados tanto do RIBO-seq quanto do RNA-seq durante a análise dos dados. O translatome investigado é definido pela normalização das pegadas ribossômicas à abundância de mRNA16. Os dados do RNA-seq também podem ajudar a identificar a clonagem ou sequenciamento de artefatos17.
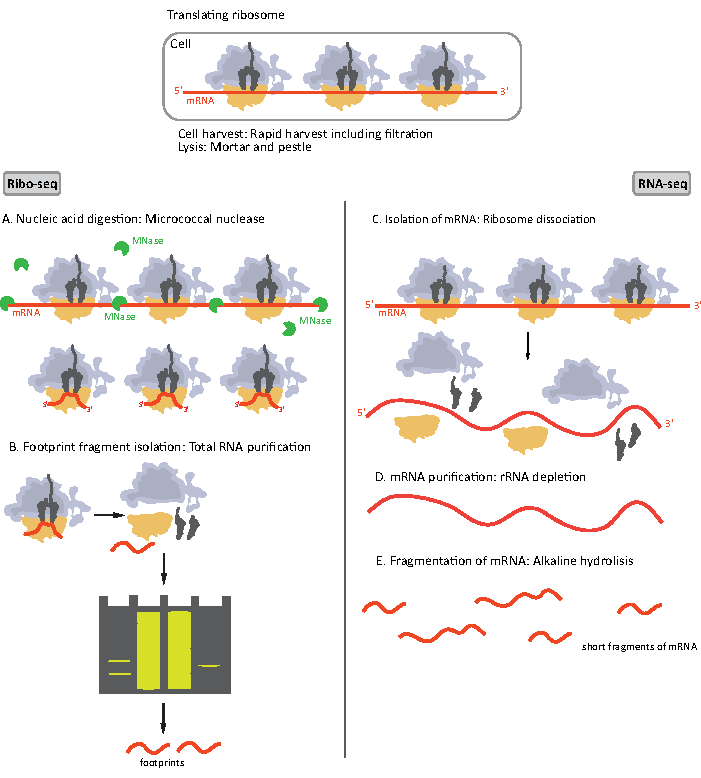
Figura 1. Esquemas de preparação de amostras de mRNA para RIBO-seq e RNA-seq. Para a preparação da biblioteca RIBO-seq, o RNA é digerido com MNase (A), seguido pela seleção de tamanho de RF de ~28-30 nt de comprimento (B); para RNA-seq RNA é isolado (C), esgotado de rRNA (D), e o mRNA resultante é aleatoriamente fragmentado em fragmentos de comprimentos variados (E). Clique aqui para ver uma versão maior desta figura.
As etapas iniciais do procedimento de preparação da amostra para RIBO-seq e RNA-seq diferem ligeiramente(Figura 1). Para o perfil ribossômico, o liseto precisa ser digerido por uma endonuclease específica para degradar as moléculas de mRNA não protegidas pelos ribossomos. Nos protocolos padrão, os monossomos obtidos são recuperados por uma ultracentrifugação de almofada de sacarose ou uma ultracentrifugação de gradiente de sacarose8,14. Neste artigo, mostramos que esta etapa não é necessária para isolar a RF necessária para o RIBO-seq em bactérias, da mesma forma para as células eucarióticas18, e que a seleção de tamanho dos fragmentos de comprimento apropriado do gel de poliacrilamida é suficiente.
Para RNA-seq, o mRNA é obtido pelo esgotamento do rRNA do total de moléculas de RNA – rRNA hibridizem-se às sondas oligonucleotidas biotinínas que se ligam às contas magnéticas revestidas de streptavidin. Os complexos rRNA-oligonucleotídeo-contas são então removidos da amostra com um ímã resultando em uma amostra esgotada de rRNA19,20. As moléculas de mRNA purificadas são então fragmentadas aleatoriamente por hidrólise alcalina. Os fragmentos obtidos de mRNA, bem como as pegadas ribossômicas são convertidos em bibliotecas cDNA e preparados para sequenciamento profundo(Figura 2). Isso envolve o reparo necessário após a hidrólise alcalina (para mRNA) e digestão enzimática (para RF): desfosforilação de extremidades de 3′ seguida de fosforilação de extremidades de 5′. Os próximos passos são a ligadura dos adaptadores e a transcrição reversa para criar inserções cDNA emolduradas por sequências necessárias para o sequenciamento de próxima geração (NGS) usando a plataforma Illumina. A última fase da preparação da biblioteca é uma reação pcr na qual os construtos são amplificados e rotulados com códigos de barras específicos de amostra para permitir multiplexagem e sequenciamento de várias amostras em um único canal. Antes do sequenciamento, a qualidade e a quantidade das bibliotecas são avaliadas pelo DNA de alta sensibilidade na eletroforese do chip. bibliotecas cDNA com parâmetros apropriados podem então ser agrupadas e sequenciadas. O sequenciamento pode ser realizado em diferentes plataformas de Illumina, como MiSeq, NextSeq ou HighSeq, dependendo do número de bibliotecas, comprimento de leitura necessário e profundidade de sequenciamento. Após o sequenciamento, a análise bioinformática é realizada.

Figura 2. Preparação da biblioteca. A preparação da biblioteca inclui o reparo de extremidades, ligadura dos adaptadores, transcrição reversa e amplificação com codificação de barras. Clique aqui para ver uma versão maior desta figura.
O perfil ribossomo é um método universal que pode ser facilmente modificado e ajustado de acordo com a questão científica. Originalmente foi usado na levedura1, mas logo depois foi aplicado às células bacterianas21, bem como organismos modelo eucarióticos, incluindo camundongos10,zebrafish22, mosca de frutas23 e Arabidopsis thaliana24. Também foi utilizado para o estudo de diferentes tipos ribossósmicos: citoplasmásmico, mitocondrial25,26 e cloroplasto27,28. Em eucariotes RIBO-seq é comumente adaptado e refinado para investigar aspectos específicos da tradução, incluindo iniciação10,11,29,30,31,32, alongamento1,10,11,31,33, ribossomo parando33 e alteração de conformação33. A maioria das modificações envolve o uso de diferentes inibidores de tradução. No entanto, nas bactérias, estudos análogos têm sido difíceis de realizar devido à escassez de inibidores com o mecanismo de ação necessário34. O inibidor de tradução mais usado em bactérias é o clorofífenicol (CAM), que se liga ao centro de transferência de peptidyl (PTC) e impede o posicionamento correto do aminoacílico-tRNA no local A. Como resultado, cam impede a formação de um vínculo peptídeo que leva à prisão dos ribossomosalongamentos 35. Outros exemplos de inibidores de tradução em bactérias são a tetraciclina (TET)36, retapamulina (RET)34 e Onc11237 que foram usados para investigar sites de iniciação de tradução. O TET, que impede a entrega de tRNA ao ribossomo, sobrepondo-se diretamente com o loop anticodon de tRNA no local A, foi originalmente aplicado para verificar os resultados obtidos do tratamento CAM, uma vez que ambos são antibióticos inibindo a alongamento da tradução38. Foi encontrado tet para detectar TIS primário, no entanto foi incapaz de revelar TIS36interno . RET se liga no PTC do ribossomo bacteriano, e impede a formação da primeira ligação de peptídeo interferindo com um alongador aminoacyl-tRNA no site A. A aplicação de RET resulta em prisão de ribossomos tanto nas TISs primárias quanto internas34. Onc112, um peptídeo antimicrobiano rico em proline, liga-se no túnel de saída e bloqueia a ligação aminoacíl-tRNA no local ribossômico A. Como resultado, o Onc112 impede que os complexos de iniciação entrem na fase de alongamento37.
A principal informação que o perfil ribossomo fornece é a densidade de ribossomos e sua posição no mRNA. Foi aplicado com sucesso para investigar a expressão genética diferencial no nível da tradução em várias condições de crescimento1,6, medir a eficiência translacional1,38,39 e detectar eventos de regulação de tradução, como a pausa ribossômica10. RIBO-seq também permite descobrir a tradução de ncRNA anotado, pseudogenes e pequenos quadros de leitura abertos não anotados (ORF) levando à identificação de genes de codificação de proteínas novos e/ou muito curtos10,12,22,30,37. Nesses casos, ribo-seq pode ajustar e melhorar a anotação do genoma. Com sua alta sensibilidade para a identificação de ORFs traduzidos e sua natureza quantitativa, o perfil ribossomo também pode servir como proxy para a determinação proteome ou para auxiliar estudos de proteômica31,34,39. Ao mapear o TIS, o perfil ribossomo revela isóformes n-terminalmente estendidos e truncados de proteínas conhecidas10,32. RIBO-seq também foi adaptado para estudar o dobrável co-translacional das proteínas14,21,24. Este método permite medir as taxas de alongamento1,10,39 ou velocidades de decodificação de códons individuais6 e ajuda no desenvolvimento de modelos quantitativos de tradução17. O método de criação de perfil ribossomo também é capaz de fornecer insights mecanicistas sobre a pausa ribossa em bactérias7,15,17, frameshifting40, stop-codon readthrough21, defeitos de terminação/reciclagem41,42 e alterações de conformação ribossômica33 em eucariotos. RIBO-seq também foi adaptado para examinar o impacto de fatores trans-acionados específicos na tradução, como miRNAs6 e proteínas de ligação de RNA em eucariotes16,43. No entanto, é importante reconhecer que o projeto experimental e a resolução obtida do RIBO-seq determinam a quantidade de informações que podem ser extraídas dos dados de sequenciamentoresultantes 12.
