Die Ribosomenprofilierungstechnik (RIBO-seq) wurde im Labor von Jonathan Weissman an der University of California, San Francisco1entwickelt. Im Vergleich zu anderen Methoden zur Untersuchung der Genexpression auf translationaler Ebene konzentriert sich RIBO-seq auf jede Ribosombindung an mRNA und liefert Informationen über seine Position und die relative Anzahl der Ribosomen auf einem Transkript. Es ermöglicht die Überwachung des Prozesses der Proteinsynthese in vivo und kann eine einzelne Codonauflösung und -genauigkeit liefern, die die Messung der Ribosomendichte sowohl auf der einzelnen mRNA als auch entlang des gesamten Transkriptoms in der Zelle ermöglicht. Grundlage der RIBO-seq-Technik ist die Tatsache, dass das Ribosom während der Translation das mRNA-Molekül bindet und so das vergrabene Fragment des Transkripts vor einer Ribonuklease-Verdauung schützt. Nach Zugabe der Ribonuklease wird die ungeschützte mRNA verdaut und die von Ribosomen eingeschlossenen Fragmente – typischerweise von ~28-30 nt lang – bleiben intakt. Diese Fragmente, die als ribosomale Fußabdrücke (RF) bezeichnet werden, können dann isoliert, sequenziert und auf das Transkript abgebildet werden, aus dem sie stammen, was zur Erkennung der genauen Position der Ribosomen führt. Tatsächlich wird die Ribosomenfähigkeit zum Schutz von mRNA-Fragmenten seit den 1960er Jahren verwendet, um ribosomale Bindungs- und Translationsinitiierungsstellen (TIS)2,3,4zu untersuchen. Mit dem Fortschritt in der Tiefensequenzierungstechnologie ist RIBO-seq jedoch zu einem Goldstandard für die Translationsüberwachunggeworden 5, der durch das Ribosomengagement genomweite Informationen über die Proteinsynthese liefern kann6. Ribosomenprofilierung füllte die technologische Lücke, die zwischen der Quantifizierung des Transkriptoms und des Proteoms bestand6.
Um ein Ribosomen-Profiling durchzuführen, müssen wir Zelllysat des Organismus erhalten, der unter den untersuchten Bedingungen gewachsen war. Die Störung dieser Bedingungen während der Zellentnahme und -lyse kann unzuverlässige Daten liefern. Um dies zu verhindern, werden häufig Translationsinhibitoren, schnelle Ernte und Schockgefrieren in flüssigem Stickstoff verwendet. Zellen können durch kryogenes Mahlen in einem mechanischen Homogenisator wie einer Mischermühle7,8 odereinem Perlenschläger9und durch Trituration durch eine Pipette10 oder mit einer Nadel11lysiert werden. Der Lysepuffer kann kurz vor oder kurz nach der Pulverisierung der Zellen zugegeben werden. In unserem Protokoll verwenden wir flüssigen Stickstoff zur Vorkühlung von Mörtel und Stößel sowie Aluminiumoxid als schonenderen Ansatz zur Störung der bakteriellen Zellwand, was eine RNA-Scherung verhindert, die häufig bei Methoden wie sonifikation auftritt. Nach der Pulverisierung fügen wir dem gekühlten Inhalt des Mörtels einen eiskalten Lysepuffer hinzu. Die Auswahl eines geeigneten Lysepuffers ist wichtig, um die beste Auflösung der ribosomalen Fußabdrücke zu erhalten. Da die Ionenstärke sowohl die HF-Größe als auch die Leserahmengenauigkeit beeinflusst, wird derzeit empfohlen, Lysepuffer mit geringer Ionenstärke und Pufferkapazität zu verwenden, auch wenn es den Anschein hat, dass die Pufferzusammensetzung die ribosomale Belegung auf mRNAs11,12nicht beeinflusst. Wichtige Bestandteile des Lysepuffers sind Magnesiumionen, deren Vorhandensein eine Dissoziation der ribosomalen Untereinheiten verhindert und spontane Konformationsveränderungen in den bakteriellen Ribosomen hemmt11,13. Calciumionen spielen ebenfalls eine bedeutende Rolle und sind essentiell für die Aktivität der Mikrokokkennuklease (MNase), die in der bakteriellen Ribosomenprofilierungsmethode verwendet wird14. Die Zugabe von Guanosin 5′-[β,γ-imido]triphosphat (GMP-PNP), einem nicht hydrolysierbaren Analogon von GTP, zusammen mit Chloramphenicol hemmt die Translation während der Lyse15.
Wenn das Lysat erhalten wird, wird es durch Zentrifugation geklärt und in zwei Portionen unterteilt, jeweils für eine RIBO-seq- und eine Hochdurchsatz-Gesamt-mRNA-Sequenzierung (RNA-seq), da sie gleichzeitig durchgeführt werden (Abbildung 1). RNA-seq bietet einen Bezugspunkt, der den Vergleich von Daten sowohl von RIBO-seq als auch von RNA-seq während der Datenanalyse ermöglicht. Das untersuchte Translatom wird durch Normalisierung ribosomaler Fußabdrücke zur mRNA-Häufigkeitdefiniert 16. Daten von RNA-seq können auch helfen, Klon- oder Sequenzierungsartefakte zu identifizieren17.
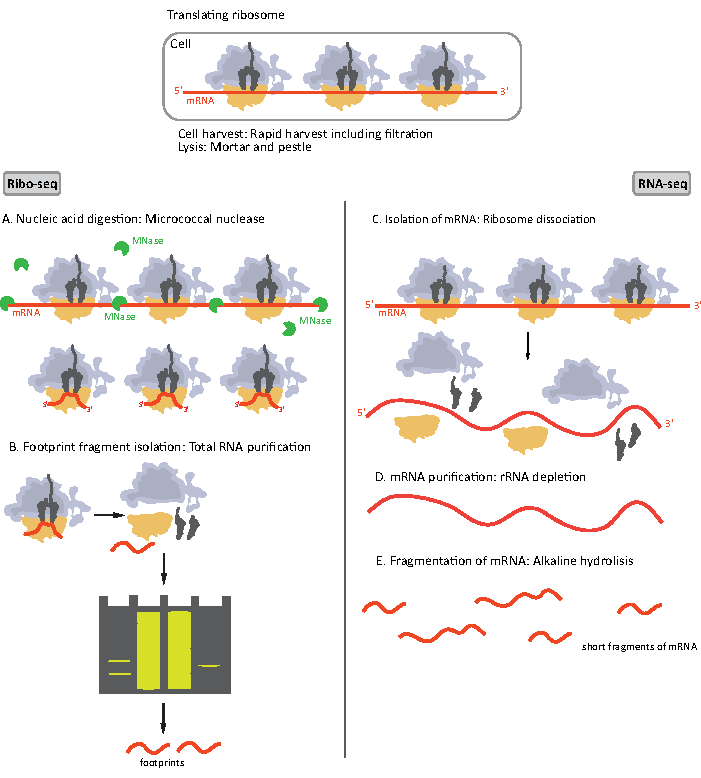
Abbildung 1. Schematische Darstellung der mRNA-Probenvorbereitung für RIBO-seq und RNA-seq. Für die RIBO-seq-Bibliotheksvorbereitung wird RNA mit MNase (A) verdaut, gefolgt von der Größenauswahl von RF von ~ 28-30 nt Länge (B); für RNA-seq wird RNA isoliert (C), von rRNA (D) erschöpft und die resultierende mRNA wird zufällig in Fragmente unterschiedlicher Länge (E) fragmentiert. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Die ersten Schritte des Verfahrens der Probenvorbereitung für RIBO-seq und RNA-seq unterscheiden sich geringfügig (Abbildung 1). Für das ribosomale Profiling muss das Lysat durch eine spezifische Endonuklease verdaut werden, um die mRNA-Moleküle abzubauen, die nicht durch die Ribosomen geschützt sind. In Standardprotokollen werden die erhaltenen Monosomen durch eine Saccharosekissen-Ultrazentrifugation oder eine Saccharosegradienten-Ultrazentrifugation8,14zurückgewonnen. In diesem Artikel zeigen wir, dass dieser Schritt nicht notwendig ist, um RF zu isolieren, die für das RIBO-seq in Bakterien erforderlich sind, ebenso für eukaryotische Zellen18, und dass die Größenauswahl der geeigneten mRNA-Fragmente aus dem Polyacrylamid-Gel ausreichend ist.
Für RNA-seq wird mRNA durch die Erschöpfung von rRNA aus der gesamten RNA erhalten – rRNA-Moleküle hybridisieren zu den biotinylierten Oligonukleotidsonden, die an die Streptavidin-beschichteten magnetischen Perlen binden. Die rRNA-Oligonukleotid-Perlenkomplexe werden dann mit einem Magneten aus der Probeentfernt,was zu einer rRNA-erschöpften Probe19,20führt. Die gereinigten mRNA-Moleküle werden dann durch alkalische Hydrolyse zufällig fragmentiert. Die erhaltenen Fragmente der mRNA sowie die ribosomalen Fußabdrücke werden in cDNA-Bibliotheken umgewandelt und für die Tiefensequenzierung vorbereitet (Abbildung 2). Dies beinhaltet die Reparatur der Enden, die nach der alkalischen Hydrolyse (für mRNA) und der enzymatischen Verdauung (für RF) erforderlich ist: Dephosphorylierung von 3′-Enden, gefolgt von Phosphorylierung von 5′-Enden. Die nächsten Schritte sind die Adapterligatur und die umgekehrte Transkription, um cDNA-Inserts zu erstellen, die von Sequenzen umrahmt werden, die für die Next Generation Sequencing (NGS) mit der Illumina-Plattform erforderlich sind. Die letzte Phase der Bibliotheksvorbereitung ist eine PCR-Reaktion, bei der die Konstrukte amplifiziert und mit probenspezifischen Barcodes markiert werden, um multiplexen und sequenzieren zu können, verschiedene Proben auf einem Kanal zu sequenzieren. Vor der Sequenzierung werden Qualität und Quantität der Bibliotheken durch die hochempfindliche DNA-On-Chip-Elektrophorese bewertet. cDNA-Bibliotheken mit entsprechenden Parametern können dann gepoolt und sequenziert werden. Die Sequenzierung kann auf verschiedenen Illumina-Plattformen wie MiSeq, NextSeq oder HighSeq durchgeführt werden, abhängig von der Anzahl der Bibliotheken, der erforderlichen Leselänge und der Sequenzierungstiefe. Nach der Sequenzierung wird die bioinformatische Analyse durchgeführt.

Abbildung 2. Bibliotheksvorbereitung. Die Bibliotheksvorbereitung umfasst die Reparatur der Enden, die Ligatur der Adapter, die umgekehrte Transkription und die Verstärkung mit Barcoding. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Die Ribosomenprofilierung ist eine universelle Methode, die sich leicht modifizieren und entsprechend der wissenschaftlichen Fragestellung anpassen lässt. Ursprünglich wurde es in Hefe1verwendet, aber kurz darauf wurde es auf Bakterienzellen21 sowie eukaryotische Modellorganismen wie Maus10,Zebrafisch22,Fruchtfliege23 und Arabidopsis thaliana24aufgetragen. Es wurde auch für die Untersuchung verschiedener Ribosomentypen verwendet: zytoplasmatische, mitochondriale25,26 und Chloroplast27,28. Bei Eukaryoten wird RIBO-seq üblicherweise angepasst und verfeinert, um spezifische Aspekte der Translation zu untersuchen, einschließlich Initiation10,11,29,30,31,32, Dehnung1,10,11,31,33, Ribosomenabwürgen33 und Konformationsänderung33. Die meisten Modifikationen beinhalten die Verwendung verschiedener Translationsinhibitoren. Bei Bakterien waren analoge Studien jedoch aufgrund des Mangels an Inhibitoren mit dem erforderlichen Wirkmechanismus schwierigdurchzuführen 34. Der am häufigsten verwendete Translationsinhibitor in Bakterien ist Chloramphenicol (CAM), das an das Peptidyltransferase-Zentrum (PTC) bindet und eine korrekte Positionierung der Aminoacyl-tRNA in der A-Stelle verhindert. Infolgedessen verhindert CAM die Bildung einer Peptidbindung, die dazu führt, dass die länglichen Ribosomen35blockiert werden. Weitere Beispiele für Translationsinhibitoren in Bakterien sind Tetracyclin (TET)36, Retapamulin (RET)34 und Onc11237, die zur Untersuchung von Translationsinitiierungsstellen verwendet wurden. TET, das die tRNA-Abgabe an das Ribosom verhindert, indem es sich direkt mit der Anticodon-Stammschleife der tRNA an der A-Stelle überschneidet, wurde ursprünglich angewendet, um die Ergebnisse der CAM-Behandlung zu überprüfen, da beide Antibiotika sind, die die Translationsdehnung hemmen38. Es wurde festgestellt, dass TET primäre TIS erkennt, konnte jedoch kein internes TIS36aufdecken. RET bindet im PTC des bakteriellen Ribosoms und verhindert die Bildung der ersten Peptidbindung, indem es mit einer Elongator-Aminoacyl-tRNA an der A-Stelle interferiert. Die Anwendung von RET führt zu einem Ribosomen-Arrest sowohl bei primären als auch bei internen TISs34. Onc112, ein prolinreiches antimikrobielles Peptid, bindet im Austrittstunnel und blockiert die Aminoacyl-tRNA-Bindung an der ribosomalen A-Stelle. Dadurch verhindert Onc112, dass Initiationskomplexe in die Dehnungsphase37eintreten.
Die Hauptinformation, die das Ribosomen-Profiling liefert, ist die Ribosomendichte und ihre Position auf der mRNA. Es wurde erfolgreich angewendet, um die differentielle Genexpression auf der Ebene der Translation in verschiedenen Wachstumsbedingungen1,6zu untersuchen, die translationale Effizienz1,38,39 zu messen und Translationsregulationsereignisse wie ribosomale Pausierung10zu erkennen. RIBO-seq ermöglicht auch die Aufdeckung der Translation von annotierter ncRNA, Pseudogenen und unangekündigten kleinen offenen Leserahmen (ORF), die zur Identifizierung neuer und/oder sehr kurzer proteinkodierender Gene10,12,22,30,37führen. In solchen Fällen kann RIBO-seq die Genomannotation verfeinern und verbessern. Mit seiner hohen Sensitivität für die Identifizierung übersetzter ORFs und seiner quantitativen Natur kann das Ribosomen-Profiling auch als Proxy für die Proteombestimmung oder zur Unterstützung von Proteomik-Studiendienen 31,34,39. Durch die Kartierung von TIS zeigt das Ribosomen-Profiling N-terminal ausgedehnte und abgeschnittene Isoformen bekannter Proteine10,32. RIBO-seq wurde auch angepasst, um die co-translationale Faltung der Proteine14,21,24zu untersuchen. Diese Methode ermöglicht die Messung von Dehnungsraten1,10,39 oder Dekodierungsgeschwindigkeiten einzelner Codons6 und hilft bei der Entwicklung quantitativer Modelle der Übersetzung17. Die Ribosomen-Profiling-Methode ist auch in der Lage, mechanistische Einblicke in die Ribosomen-Pause bei Bakterien7,15,17, Frameshifting40, Stop-Codon-Readthrough21, Abschluss- / Recyclingdefekte41,42 und ribosomale Konformationsänderungen33 in Eukaryoten zu liefern. RIBO-seq wurde auch angepasst, um den Einfluss spezifischer trans-wirkender Faktoren auf die Translation wie miRNAs6 und RNA-bindende Proteine in Eukaryoten16,43zuuntersuchen. Es ist jedoch wichtig anzuerkennen, dass das experimentelle Design und die erhaltene Auflösung von RIBO-seq die Menge an Informationen bestimmen, die aus den resultierenden Sequenzierungsdaten extrahiert werden können12.
