De niet-specifieke endocytische route van macropinocytose stelt cellen in staat om een verscheidenheid aan extracellulaire componenten, waaronder voedingsstoffen, eiwitten, antigenen en pathogenen, te internaliseren door bulkopname van extracellulaire vloeistof en zijn bestanddelen1. Hoewel belangrijk voor de biologie van talrijke celtypen, wordt in toenemende mate beschreven dat de macropinocytoseroute een essentiële rol speelt in de tumorbiologie, waar tumorcellen door macropinocytische opname in staat zijn om te overleven en zich te vermenigvuldigen in de aanwezigheid van een door voedingsstoffen uitgeputte micro-omgeving2,3. De opname van extracellulaire macromoleculen, waaronder albumine en extracellulaire matrix, en necrotisch celpuin, biedt een alternatieve voedingsbron voor biomassaproductie door aminozuren, suikers, lipiden en nucleotiden te creëren door middel van macropinosoom en lysosoom fusie-gemedieerd ladingkatabolisme4,5,6,7,8.
De inductie en regulatie van macropinocytose zijn complex en kunnen variëren afhankelijk van de cellulaire context. Tot nu toe zijn verschillende inductoren van macropinocytose geïdentificeerd en omvatten liganden, zoals epidermale groeifactor (EGF), van bloedplaatjes afgeleide groeifactor (PDGF), galectine-3 en Wnt3A9,10,11,12,13. Bovendien kunnen kweekomstandigheden die de micro-omgeving van de tumor nabootsen, activering van de route veroorzaken. Pancreas ductaal adenocarcinoom (PDAC) tumoren zijn voedingsarm, vooral voor het aminozuur glutamine, waardoor zowel kankercellen als kanker-geassocieerde fibroblasten (CAFs) afhankelijk zijn van macropinocytose voor overleving7,13,14,15. Bovendien kunnen tumorspanningen, zoals hypoxie en oxidatieve stress, deze opruimroute activeren16. Naast de vele extrinsieke beïnvloeders die macropinocytose kunnen induceren, regelen verschillende intracellulaire routes de vorming van macropinosomen. Oncogene Ras-gemedieerde transformatie is voldoende om de macropinocytische machinerie te initiëren, en meerdere kankertypen vertonen oncogene Ras-gedreven constitutieve macropinocytose4,5,9,17. Als alternatief zijn rasactiverings- en Ras-onafhankelijke routes geïdentificeerd om macropinocytose in kankercellen en CAFs10,11,15,18 te activeren. Het gebruik van verschillende in vitro modellen in combinatie met inhibitorbehandelingen heeft geresulteerd in de identificatie van verschillende macropinocytosemodulatoren, waaronder natrium-waterstofwisselaars, de kleine GTPase Rac1, fosfoinositide 3-kinase (PI3K), p21-geactiveerd kinase (Pak) en AMP-geactiveerd eiwitkinase (AMPK)4,13,15 . Gezien de veelheid aan beschreven factoren en aandoeningen die macropinocytose reguleren, is het echter denkbaar dat veel meer modulatoren en stimuli onontdekt blijven. De identificatie van nieuwe modulatoren en stimuli kan worden vergemakkelijkt door geautomatiseerde beoordeling van een groot aantal omstandigheden in een enkel experiment. Deze methodologie kan licht werpen op de factoren die betrokken zijn bij de vorming van macropinosomen en kan de ontdekking van nieuwe kleine moleculen of biologische geneesmiddelen mogelijk maken die zich op deze route richten.
Hier hebben we ons eerder vastgestelde protocol voor het bepalen van de mate van macropinocytose in kankercellen in vitro aangepast naar een 96-well microplaatformaat en geautomatiseerde beeldvorming en kwantificering19,20. Dit protocol is gebaseerd op fluorescerende microscopie, die een standaard is geworden in het veld om macropinocytose in vitro en in vivo te bepalen4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Macropinosomen kunnen worden onderscheiden van andere endocytische routes door hun vermogen om grote macromoleculen te internaliseren, zoals dextran met een hoog molecuulgewicht (d.w.z. 70 kDa)2,3,4,20,21,22,23. Macropinosomen kunnen dus worden gedefinieerd door opname van extracellulair toegediende fluorofoor-gelabelde 70 kDa dextran. Als gevolg hiervan manifesteren macropinocytische blaasjes zich als intracellulaire clusters van fluorescerende puncta met afmetingen variërend van 0,2-5 μm. Deze puncta kunnen microscopisch in beeld worden gebracht en vervolgens worden gekwantificeerd om de mate van macropinocytose in de cel te bepalen – ‘de macropinocytische index’.
In dit protocol worden de essentiële stappen beschreven om macropinosomen in aanhangende cellen in vitro te visualiseren op een 96-well microplaat en coverslips met behulp van standaard laboratoriumapparatuur (figuur 1). Daarnaast worden de aanwijzingen gegeven om de beeldacquisitie en kwantificering van de macropinocytische index te automatiseren met behulp van een multi-mode plaatlezer voor celbeeldvorming. Deze automatisering vermindert tijd, kosten en moeite in vergelijking met onze eerder beschreven protocollen19,20. Bovendien voorkomt het onbedoeld bevooroordeelde beeldvormingsacquisitie en -analyse en verbetert het daardoor de reproduceerbaarheid en betrouwbaarheid. Deze methode kan eenvoudig worden aangepast aan verschillende celtypen of plaatlezers of worden gebruikt om alternatieve macropsoomkenmerken te bepalen, zoals grootte, aantal en locatie. De hierin beschreven methode is vooral geschikt voor de screening van celkweekomstandigheden die macropinocytose induceren, de identificatie van nieuwe modulatoren of optimalisatie van medicijnconcentraties van bekende remmers.
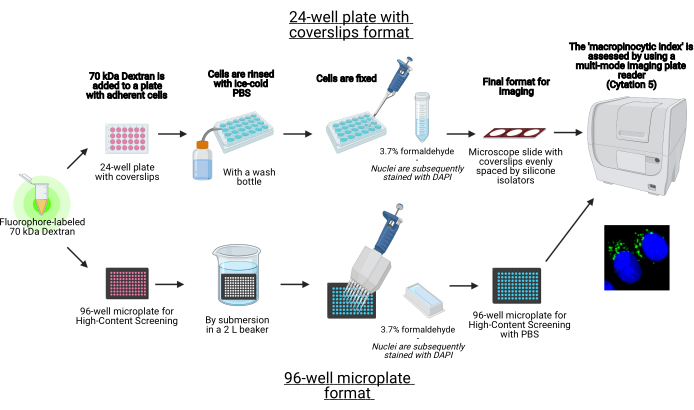
Figuur 1: Schema van de geautomatiseerde assay om de ‘macropinocytische index’ in adherente cellen te bepalen. Gemaakt met BioRender. Klik hier om een grotere versie van deze figuur te bekijken.
