Der unspezifische endozytäre Weg der Makropinozytose ermöglicht es Zellen, eine Vielzahl von extrazellulären Komponenten, einschließlich Nährstoffen, Proteinen, Antigenen und Krankheitserregern, durch Massenaufnahme von extrazellulärer Flüssigkeit und ihren Bestandteilen zu internalisieren1. Obwohl wichtig für die Biologie zahlreicher Zelltypen, wird zunehmend beschrieben, dass der Makropinozytoseweg eine wesentliche Rolle in der Tumorbiologie spielt, wo Tumorzellen durch makropinozytäre Aufnahme in der Lage sind, in Gegenwart einer nährstoffarmen Mikroumgebung zu überleben und sich zu vermehren2,3. Die Aufnahme extrazellulärer Makromoleküle, einschließlich Albumin und extrazellulärer Matrix, sowie nekrotischer Zelltrümmer bietet eine alternative Nährstoffquelle für die Biomasseproduktion, indem Aminosäuren, Zucker, Lipide und Nukleotide durch Makropinosomen- und Lysosomenfusions-vermittelte Frachtkatabolismus erzeugt werden4,5,6,7,8.
Die Induktion und Regulation der Makropinozytose ist komplex und kann je nach zellulärem Kontext variieren. Bisher wurden mehrere Induktoren der Makropinozytose identifiziert und umfassen Liganden wie den epidermalen Wachstumsfaktor (EGF), den von Thrombozyten abgeleiteten Wachstumsfaktor (PDGF), Galectin-3 und Wnt3A9,10,11,12,13. Darüber hinaus können Kultivierungsbedingungen, die die Tumormikroumgebung nachahmen, die Aktivierung des Signalwegs auslösen. Tumoren des duktalen Adenokarzinoms (PDAC) der Bauchspeicheldrüse sind nährstoffarm, insbesondere für die Aminosäure Glutamin, die dazu führt, dass sowohl Krebszellen als auch krebsassoziierte Fibroblasten (CAFs) auf Makropinozytose angewiesen sind7,13,14,15. Darüber hinaus können Tumorbelastungen wie Hypoxie und oxidativer Stress diesen Fressweg aktivieren16. Zusätzlich zu den zahlreichen extrinsischen Einflussfaktoren, die Makropinozytose induzieren können, steuern eine Vielzahl von intrazellulären Signalwegen die Bildung von Makropinosomen. Die onkogene Ras-vermittelte Transformation reicht aus, um die makropinozytische Maschinerie zu initiieren, und mehrere Krebsarten weisen eine onkogene Ras-gesteuerte konstitutive Makropinozytose auf4,5,9,17. Alternativ wurden Wildtyp-Ras-Aktivierung und Ras-unabhängige Signalwege identifiziert, um makropinozytose in Krebszellen und CAFs zu aktivieren10,11,15,18. Die Verwendung verschiedener In-vitro-Modelle in Kombination mit Inhibitorbehandlungen hat zur Identifizierung mehrerer Makropinozytosemodulatoren geführt, darunter Natrium-Wasserstoff-Austauscher, die kleine GTPase Rac1, die Phosphoinositid-3-Kinase (PI3K), die p21-aktivierte Kinase (Pak) und die AMP-aktivierte Proteinkinase (AMPK)4,13,15 . Angesichts der Vielzahl der beschriebenen Faktoren und Bedingungen, die die Makropinozytose regulieren, ist es jedoch denkbar, dass noch viel mehr Modulatoren und Reize unentdeckt bleiben. Die Identifizierung neuartiger Modulatoren und Stimuli kann durch die automatisierte Bewertung einer Vielzahl von Bedingungen in einem einzigen Experiment erleichtert werden. Diese Methodik kann Aufschluss über die Faktoren geben, die an der Bildung von Makropinosomen beteiligt sind, und kann die Entdeckung neuartiger kleiner Moleküle oder Biologika ermöglichen, die auf diesen Signalweg abzielen.
Hier haben wir unser bisher etabliertes Protokoll zur Bestimmung des Ausmaßes der Makropinozytose in Krebszellen in vitro auf ein 96-Well-Mikroplattenformat und automatisierte Bildgebung und Quantifizierung angepasst19,20. Dieses Protokoll basiert auf der Fluoreszenzmikroskopie, die zu einem Standard auf dem Gebiet der Bestimmung der Makropinozytose in vitro und in vivo geworden ist4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Makropinosomen können von anderen endozytären Signalwegen durch ihre Fähigkeit unterschieden werden, große Makromoleküle wie hochmolekulares Dextran (d.h. 70 kDa)2,3,4,20,21,22,23 zu internalisieren. Somit können Makropinosomen durch aufnahme von extrazellulär verabreichtem Fluorophor-markiertem 70 kDa Dextran definiert werden. Infolgedessen manifestieren sich makropinozytäre Vesikel als intrazelluläre Cluster fluoreszierender Punktika mit Größen von 0,2-5 μm. Diese Puncta können mikroskopisch abgebildet und anschließend quantifiziert werden, um das Ausmaß der Makropinozytose in der Zelle – den “makropinozytären Index” – zu bestimmen.
In diesem Protokoll werden die wesentlichen Schritte zur Visualisierung von Makropinosomen in adhärenten Zellen in vitro auf einer 96-Well-Mikroplatte und Deckgläsern mit Standardlaborgeräten beschrieben (Abbildung 1). Darüber hinaus werden die Anweisungen zur Automatisierung der Bildaufnahme und Quantifizierung des makropinozytären Index mit einem cell imaging Multimode-Plattenleser bereitgestellt. Diese Automatisierung reduziert Zeit, Kosten und Aufwand im Vergleich zu unseren zuvor beschriebenen Protokollen19,20. Darüber hinaus vermeidet es eine unbeabsichtigt verzerrte Bilderfassung und -analyse und erhöht dadurch die Reproduzierbarkeit und Zuverlässigkeit. Diese Methode kann leicht an verschiedene Zelltypen oder Plattenleser angepasst oder zur Bestimmung alternativer Makropinosomenmerkmale wie Größe, Anzahl und Position verwendet werden. Das hierin beschriebene Verfahren eignet sich insbesondere für das Screening von Zellkulturbedingungen, die makropinozytose induzieren, die Identifizierung neuartiger Modulatoren oder die Optimierung von Wirkstoffkonzentrationen bekannter Inhibitoren.
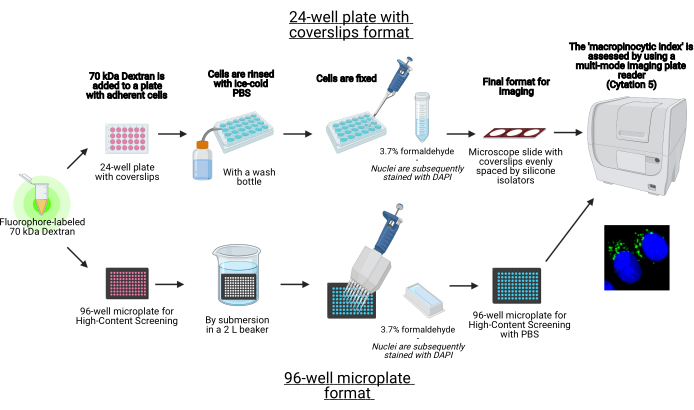
Abbildung 1: Schematische Darstellung des automatisierten Assays zur Bestimmung des “makropinozytären Index” in adhärenten Zellen. Erstellt mit BioRender. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
