Митохондрии выполняют ключевое обеспечение энергией и представляют собой компартментированную органеллу, способствующую существенным клеточным биоэнергетическим и метаболическим процессам, таким как анаболизм нуклеотидов, липидов и аминокислот, биогенез железо-серного кластера и участвуют в передаче сигналов, таких как контролируемая гибель клеток 1,2,3 . Митохондриальная биоэнергетика посредством окислительного фосфорилирования способствует практически всем клеточным процессам внутри клетки, и, следовательно, митохондриальные дисфункции первичного или вторичного происхождения связаны с широким спектром заболеваний 4,5. Митохондриальная дисфункция включает в себя не только изменения в структуре или плотности митохондрий, но и в качестве и регуляции дыхательной системы6. Этот качественный элемент включает в себя управление субстратом, характеристики связи, посттрансляционные модификации, динамику крист и дыхательные суперкомплексы 7,8. Поэтому точный анализ митохондриальной биоэнергетики для экспериментальных и диагностических подходов к оценке энергетического обмена клетки важен для здоровья и болезней.
Митохондриальное окислительное фосфорилирование (OXPHOS) представляет собой последовательность реакций в дыхательной системе или системе переноса электронов (ETS) для генерации клеточной энергии через аденозинтрифосфат (АТФ)9. Мультиферментативная стадия для использования энергии от потока электронов через комплексы I и II к комплексу IV генерирует электрохимический протонный градиент через внутреннюю митохондриальную мембрану, впоследствии используемый для фосфорилирования аденозиндифосфата (АДФ) до АТФ через комплекс V (F1FO АТФ-синтаза) (Рисунок 1А).
Во-первых, два электронных носителя образуются во время трикарбоксового цикла (TCA), гликолиза и пируватного окисления: никотинамидадениндинуклеотид (NADH) и дигидрофлавинадениндинуклеотид (FADH2). NADH окисляется в комплексе I (NADH дегидрогеназа), в ходе которого два электрона переносятся в коэнзим Q (хинон восстанавливается до хинола), в то время как протоны закачиваются в межмембранное пространство (IMS). Во-вторых, комплекс II (сукцинатдегидрогеназа) окисляет FADH2 и подает электроны в коэнзим Q без перекачки протонов. В-третьих, в комплексе III (цитохром c оксидоредуктаза) электроны из коэнзима Q переносятся в цитохром c, в то время как протоны закачиваются в IMS. В-четвертых, цитохром c переносит электроны в комплекс IV (цитохром c оксидаза), конечный комплекс для перекачки протонов, и где кислород функционирует как акцептор электронов для ассимиляции протонов, в конечном итоге образуя воду. Именно этот кислород потребляют митохондрии, который можно измерить с помощью оксиграфа. Наконец, протоны, полученные из комплекса I, комплекса III и комплекса IV, используются для вращения комплекса V, тем самым генерируя АТФ9.
Важно отметить, что перенос электронов происходит не только линейным образом, иначе обозначаемым как цепь переноса электронов. Вместо этого электроны могут быть перенесены в пул коэнзима Q через несколько дыхательных путей и облегчают конвергентный поток электронов. NADH-субстраты и сукцинат, например, могут входить через комплекс I и комплекс II соответственно. Электроны от окисления жирных кислот могут быть пожертвованы через электрон-переносящий флавопротеиновый комплекс. Действительно, всесторонний анализ OXPHOS требует целостного подхода с соответствующими топливными субстратами (рисунок 1A).
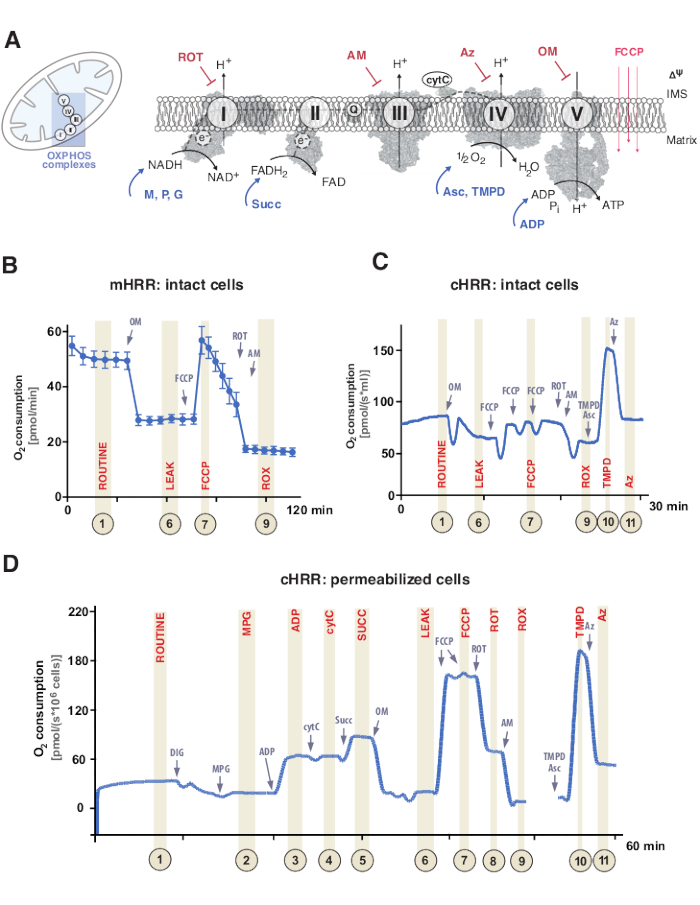
Рисунок 1: Митохондриальное окислительное фосфорилирование и специфические протоколы субстрата и ингибитора. (А) Митохондрии и схема системы переноса электронов (CI-CIV) и митохондриальная F1F0 АТФ-синтаза (CV). Все структуры из PDB. На рисунках изображены только субстраты и ингибиторы, описанные в этом исследовании). (B) Образец следа потока кислорода в неповрежденных ячейках HEK293 с использованием стандартного протокола в устройстве mHRR. (C) Образец следа потока кислорода в неповрежденных ячейках HEK293 с использованием стандартного протокола в устройстве cHRR. (D) Образец следа потока кислорода в пермеабилизированных фибробластах человека от здорового донора с соответствующим протоколом SUIT. Сокращения: 1 = Рутинное дыхание неповрежденных клеток; 2 = состояние 2; 3 = государство 3(I); 4 = состояние 3(I) с cytC; 5 = состояние 3 (I+II); 6 = Утечка(ОМ); 7 = емкость ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Ротенон, AM = Антимицин, АТФ = Аденозинтрифосфат, Az = Азид, OM = Олигомицин, FCCP = Карбонилцианид-трифтор-метоксифенилгидразон; Asc = Аскорбат, TMPD = N,N,N’,N’-тетраметил-p-фенилендиамин, Succ = Сукцинат, M = Малат, P = Пируват, ADP = Аденозиндифосфат, NAD = Никотинамиддениндинуклеотид, IMS = Межмембранное пространство, FAD = Флавинадениндинуклеотид. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Анализ емкости митохондриального OXPHOS с использованием HRR стал инструментальным биохимическим методом диагностической ценности не только для первичных митохондриальных дефектов10,11, но и для всех других областей биологии, таких как рак и старение12 лет. HRR позволяет определить клеточное дыхание путем анализа митохондриальной емкости OXPHOS, которая непосредственно отражает индивидуальную или комбинированную недостаточность митохондриального дыхательного комплекса и косвенно связана с клеточной дисфункцией и измененным энергетическим метаболизмом9. Методологически измерения дыхания выполняются с использованием клеток, тканей или изолированных митохондрий 11,13,14, с замороженным материалом, только частично подходящим15,16. Показано, что замороженная ткань имеет неповрежденную ETS с сохраненной суперкомплексной стабильностью15. Таким образом, в отличие от традиционных промежуточных продуктов TCA, соответствующие субстраты непосредственно подаются в ETS. Однако связь между ETS и синтезом АТФ теряется, поскольку целостность мембраны нарушается из-за повреждения замораживанием (образование кристаллов льда).
Эксперименты по дыханию обычно проводятся при физиологической температуре 37 °C для эндотерм в непроницаемых или пермеабилизированных клетках или тканях. В то время как первый рассматривает цитозольный метаболический контекст, последний обеспечивает энергетический вклад отдельных комплексов OXPHOS и АТФазы путем добавления специфических субстратов (и ингибиторов). Последовательность и вариация субстратов и ингибиторов привели к разработке разнообразного массива протоколовSUIT 17 и анализов18 для решения различных научных вопросов функции OXPHOS (рассмотрено в разделе12). Основной протокол клеточного дыхания оценивает четыре различных состояния: i) рутинное дыхание – дыхание в соответствующей дыхательной среде без какого-либо добавления субстратов или ингибиторов, потребляющих только эндогенные субстраты. Это состояние может выявить общий OXPHOS или вторично-индуцированные дефекты дыхания, вызванные, например, измененными профилями метаболитов. Далее добавление ингибитора АТФазы олигомицина выявляет проницаемость внутренней митохондриальной мембраны к протонам, определяемую как ii) утечка дыхания. Последующее титрование протонофора, такого как разъединитель карбонилцианида-трифтор-метоксифенилгидразон (FCCP), позволяет определить состояние, при котором емкость ETS максимальна в режиме открыто-трансмембранного протонного контура, определяемом как iii) несвязанное дыхание. Важно отметить, что несвязанное состояние может также возникать при экспериментальных вмешательствах из-за чрезмерного механического повреждения митохондриальных мембран. И наоборот, несвязанное состояние относится к дыхательному разъединению внутренним механизмом, который физиологически контролируется. Наконец, полное ингибирование ИТС путем добавления комплекса III ингибитора антимицина и комплекса I ингибитора ротенона определяет остаточное потребление кислорода (ROX) от немитохондриальных процессов, потребляющих кислород (Рисунок 1A-C).
Митохондриальная биоэнергетика состоит из пяти различных состояний дыхания19,20. Дыхание в состоянии 1 не имеет каких-либо дополнительных субстратов или АДФ, за исключением того, что эндогенно доступно. После добавления АДФ, но все же, без субстратов, достигается дыхание состояния 2. Когда добавляются подложки, позволяющие переносить электроны и синтез АТФ, достигается дыхание состояния 3. В этом состоянии емкость OXPHOS может быть определена при насыщенных концентрациях АДФ, неорганических фосфатов, кислорода, NADH- и сукцинат-связанных субстратов. Состояние дыхания 4 или дыхание LEAK может быть определено как состояние без АДФ или химически ингибированных АТФ-синтаз при наличии достаточного количества субстратов. Наконец, когда весь кислород обеднен (бескислородный) в условиях закрытой камеры, наблюдается дыхание в состоянии 5.
Существует несколько методов оценки клеточных энергетических состояний14 с двумя устройствами, доминирующими в текущей оценке OXPHOS в реальном времени путем анализа потребления кислорода, измеренного как функция снижения кислорода с течением времени в системе с закрытой камерой с различной применимостью в зависимости от экспериментальной модели и исследовательского вопроса: респирометр высокого разрешения Oroboros 2k и внеклеточный анализатор потока Seahorse XF. Оба прибора регистрируют скорость потребления кислорода как снижение пикомолей (пмоль) кислорода (О2) в секунду как абсолютное значение в камере или микропластине скважины. Удельное потребление кислорода на массу получают путем нормализации соответствующего потребления кислорода в определенном буферном рецепте на количество клеток (миллионы), вес ткани (мг) или количество белка.
O2k (Oroboros Instruments) представляет собой закрытую двухкамерную систему, оснащенную полярографическим датчиком кислорода (сокращенно камерный респирометр высокого разрешения: cHRR). Каждая экспериментальная камера вмещает 2 мл жидкости, которая поддерживается однородной магнитными мешалками. Полярографический датчик кислорода использует амперометрический подход для измерения кислорода: он содержит катод золота, анод хлорида серебра / серебра, а между раствором KCI создается электрохимическая ячейка, на которую подается напряжение (0,8 В). Кислород из пробирной среды диффундирует через 25-мкм фторированную этиленпропиленовую мембрану (O 2-проницаемую) и подвергается восстановлению на катоде, производя перекись водорода. На аноде серебро окисляется перекисью водорода, генерируя электрический ток. Этот электрический ток (ампер) линейно связан с парциальным давлением кислорода. Парциальное давление кислорода и коэффициент растворимости кислорода пробирной среды используются для расчета концентрации кислорода. Поскольку парциальное давление кислорода зависит от экспериментальной температуры, а полярографические измерения чувствительны к температуре, колебания температуры нуждаются в точном (±0,002 °C) регулировании нагревательным блоком Пельтье. Температура может контролироваться в диапазоне 4 °C и 47 °C.
Внеклеточный анализатор потока Seahorse XF (Agilent) представляет собой пластинчатую систему с 24- или 96-луночным форматом микропластин, в которой три флуоресцентных электрода измеряют потребление кислорода с течением времени в каждой скважине (сокращенно респирометр высокого разрешения на основе микропластин: mHRR). Максимум четыре порта в картридже для анализа доступны для автоматического впрыска во время анализа. Анализ содержит несколько циклов, каждый из которых состоит из трех фаз: 1) смешивание, 2) ожидание и 3) измерение. Во время фазы измерения датчики опускаются в микропластинку, создавая временно закрытую камеру, содержащую объем 7-10 мкл для измерения излучаемого света. Этот свет излучается встроенными в полимер флуорофорами на кончике сенсорных зондов, которые ощущают O2 на основе закалки фосфоресценции. Интенсивность флуоресцентного сигнала пропорциональна O2 и зависит от температуры датчика и пробирной среды. Поэтому точная оценка кислорода требует относительного подхода с фоновой скважиной без какой-либо выборки. Восстановление концентрации кислорода происходит во время фазы перемешивания, когда датчик перемещается вверх и вниз, чтобы смешать объем над временной камерой. Каждый цикл вычисляет одну норму потребления кислорода. Температура может контролироваться в диапазоне 16 °C и 42 °C.
HRR является золотым стандартом для оценки клеточной биоэнергетики при первичных и митохондриальных заболеваниях и общем клеточном метаболизме. В этом исследовании представлены основные протоколы HRR для оценки функции OXPHOS в клетках и тканях.
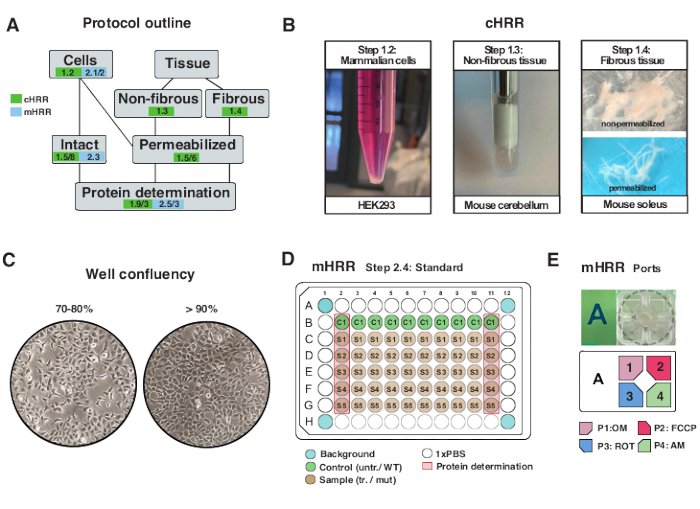
Рисунок 2: Рабочий процесс для клеточных и тканевых препаратов для cHRR и клеточной подготовки к mHRR респирометрии. (A) Краткое описание предоставленных протоколов. (B) Клетки млекопитающих (этап 1.2): гранулы HEK293, равные 3 x 106 клеткам (левая панель). Неволокнистая ткань (этап 1.3): Получение лизата мозжечка мышей в 2 мл тефлонового гончара (средняя панель). Сапонин-индуцированная пермеабилизация скелетных мышц (шаг 1.4) правая панель) для cHRR-респирометрии. (C) Стандартная схема посева микропластин (этап 2.4) и проверка слияния для анализа эукариотических клеток (HEK293) для респирометрии mHRR. (Д, Д) Схема загрузки инжекторного порта для респирометрии mHRR (шаг 2.4). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
