La formación de hueso por el osteoblasto se acompaña de destrucción coordinada o reabsorción de huesos por osteoclastos. El equilibrio entre la formación de hueso osteoblástico y la reabsorción de osteoclastos es un proceso acoplado que describe el recambio óseo o la remodelación, que es esencial para la homeostasis esquelética. La disfunción osteoblástica conduce a una formación ósea deteriorada y da lugar a diversas enfermedades, incluida la osteoporosis 1,2,3. La diferenciación ex vivo/in vitro de células madre del estroma de la médula ósea (BMSC) a precursores de osteoblastos y osteoblastos maduros da como resultado la formación y deposición de la matriz ósea mineralizada en el vaso de cultivo a lo largo del tiempo 4,5,6. Esta formación ósea por el osteoblasto requiere una cantidad significativa de energía celular. Específicamente, se ha demostrado que la síntesis y secreción de colágeno dependen en gran medida de las proporciones celulares de ATP: ADP, y presumiblemente, el tráfico y la secreción de vesículas mineralizadas requieren ATPadicional 7,8,9,10,11. Muchos investigadores han demostrado que el proceso de osteoblastogénesis y función osteoblástica requiere un suministro adecuado de energía para satisfacer la demanda metabólica de formación ósea 12,13,14,15,16. Por lo tanto, el objetivo de este método es caracterizar el estado bioenergético de las células del estroma murino primario a lo largo de la diferenciación de osteoblastos utilizando el analizador de flujo metabólico celular en tiempo real. Estas técnicas ayudan a desarrollar una mejor comprensión de la homeostasis esquelética, lo que en última instancia puede conducir al desarrollo de nuevas opciones terapéuticas capaces de mejorar los trastornos esqueléticos.
El analizador de flujo metabólico celular en tiempo real se puede utilizar para medir la tasa de consumo de oxígeno (OCR) y la tasa de acidificación extracelular (ECAR) de los osteoblastos vivos, que corresponde a la producción de ATP mitocondrial y glicolítico, respectivamente. Fundamental para esta metodología es el hecho de que un ion H+ por lactato se libera durante la glucólisis en la conversión de glucosa en lactato, lo que altera el pH de los medios reflejado en los valores de ECAR. Por el contrario, durante el ciclo del TCA (ácido tricarboxílico), la fosforilación oxidativa a través de las mitocondrias produce CO2 utilizando o consumiendo oxígeno, y por lo tanto el monitoreo de OCR refleja este proceso metabólico. El analizador mide tanto OCR como ECAR en el microambiente extracelular simultáneamente y en tiempo real, lo que permite un tremendo potencial al estudiar la bioenergética celular 6,17. Además, realizar estos ensayos es relativamente sencillo y fácilmente personalizable dependiendo del objetivo experimental. Se han empleado técnicas similares para comprender mejor la regulación metabólica de las células T del sistema inmunitario18,19, el inicio del cáncer y la progresión20, junto con otros múltiples tipos de células que contribuyen a los síndromes metabólicos21,22.
Las ventajas del analizador de flujo metabólico en tiempo real sobre las técnicas alternativas incluyen (1) la capacidad de medir la bioenergética celular de las células vivas en tiempo real, (2) la capacidad de realizar ensayos con un número relativamente pequeño de células (requiere tan solo 5.000 células), (3) los puertos de inyección para manipular paralelamente múltiples tratamientos en un sistema de 96 pocillos de alto rendimiento, (4) el uso de imágenes celulares automatizadas sin etiquetas radiactivas para la normalización18, 23,24. Los siguientes métodos tienen como objetivo proporcionar una descripción generalizada pero detallada del monitoreo de la bioenergética celular en BMSC murinas a lo largo de la diferenciación de osteoblastos utilizando el analizador. Incluirá ensayos realizados rutinariamente; sin embargo, al igual que con muchas técnicas y métodos, se recomienda encarecidamente que los laboratorios individuales determinen detalles específicos para sus experimentos.
Selección de ensayos y diferentes tipos de ensayos disponibles: Una amplia variedad de kits de ensayo y reactivos están disponibles para estudiar la bioenergética de las células, al tiempo que se garantiza la fiabilidad y consistencia de los resultados experimentales. Además, el software de escritorio también ofrece plantillas de ensayo que se pueden personalizar fácilmente. El ensayo se puede definir en función de las necesidades del usuario para medir diferentes parámetros metabólicos. Estos ensayos pueden ser modificados de varias maneras en función del objetivo experimental y/o la cuestión científica. Por ejemplo, con cuatro puertos de inyección, se pueden inyectar múltiples compuestos en los medios de ensayo para analizar la respuesta celular específica de cada vía metabólica.
Prueba de fenotipo de energía celular: Este ensayo mide el fenotipo metabólico y el potencial metabólico de las células vivas. Este ensayo también se recomienda como el primer paso para tener una idea generalizada del metabolismo específico de la vía. Se inyecta una mezcla de oligomicina A-un inhibidor de la ATP sintasa y cianuro de carbonilo 4-(trifluorometoxi) fenilhidrazona (FCCP), un agente de desacoplamiento mitocondrial para comprender el potencial de energía celular. La inyección de oligomicina A inhibe la síntesis de ATP, lo que resulta en un aumento en la tasa de glucólisis (ECAR) para permitir que las células satisfagan sus demandas de energía; por otro lado, la inyección de FCCP resulta en un mayor OCR debido a la despolarización de la membrana mitocondrial. Esencialmente, este ensayo representa la respiración metabólica basal, y después de las inyecciones duales, empujones o tensiones, la respuesta metabólica. Sobre la base de estos parámetros, el software traza OCR y ECAR de las células clasificando las células como estado aeróbico, quiescente, glucolítico o energético a lo largo del tiempo25,26.
Ensayo de tasa de producción en tiempo real de ATP: Esto mide la producción celular de ATP simultáneamente a partir de la glucólisis y la respiración mitocondrial. Este ensayo mide cuantitativamente los cambios metabólicos de las dos vías de energía y proporciona datos sobre las tasas de producción de ATP mitocondrial y glicolítico a lo largo del tiempo. El ensayo obtiene datos basales de OCR y ECAR seguidos del cálculo de la tasa de producción de ATP mitocondrial a través de la inyección de oligomicina A y la tasa de producción de ATP glicolítico a través de la inyección de rotenona + mezcla de antimicina A (inhibición total de la función mitocondrial), lo que resulta en acidificación mitocondrial17,27.
Prueba de esfuerzo de mitocondrias celulares (o prueba de esfuerzo de mito celular): Esto mide la función mitocondrial a través de la respiración ligada al ATP, cuantifica la bioenergética celular, identifica la disfunción mitocondrial y mide la respuesta de las células al estrés. Varios parámetros, incluida la capacidad respiratoria basal y sobrante, la respiración ligada a ATP, la respiración máxima y el consumo de oxígeno no mitocondrial, se pueden obtener en un solo ensayo. Este ensayo implica inyecciones secuenciales de oligomicina A, FCCP (agente desacoplamiento mitocondrial), una mezcla de inhibidores de rotenona /antimicina A para analizar eficientemente el efecto de estos en la función mitocondrial28.
Prueba de flexibilidad de flexión de combustible mito: Esto mide la tasa de respiración mitocondrial por la oxidación de los tres combustibles mitocondriales primarios por la presencia y ausencia de sus inhibidores. La inhibición secuencial de glucosa, glutamina y ácidos grasos ayuda a medir la dependencia, capacidad y flexibilidad de las células y la dependencia de las células en varias vías celulares para satisfacer la demanda de energía. Cuando las mitocondrias no pueden satisfacer las demandas de la vía bloqueada de interés oxidando otros combustibles, las células entran en un estado de dependencia. La capacidad de las células se calcula mediante la inhibición de las otras dos vías alternativas seguida de la inhibición de la vía de interés. La flexibilidad de las células ayuda a comprender la capacidad de las mitocondrias para compensar y satisfacer las necesidades de combustible de la vía inhibida. Se calcula restando la dependencia de las células de la capacidad de las células. Tres inhibidores diferentes se utilizan de forma independiente o como una mezcla de dos para calcular eficazmente los parámetros del ensayo. El ácido 2-ciano-3-(1-fenil-1H-indol-3-il)-2-propenoico (UK5099) inhibe la oxidación de la glucosa al bloquear el portador de piruvato en la glucólisis. El sulfuro de etilo bis-2-(5-fenilacetamidado-1,3,4-tiadiazol-2-il) (BPTES) inhibe la vía de oxidación de la glutamina, y el etomoxir inhibe la oxidación de los ácidos grasos de cadena larga29.
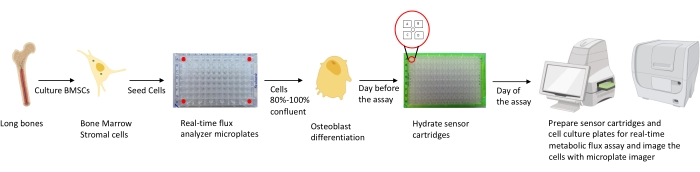
Figura 1: Representación esquemática de la metodología de cultivo y preparación de osteoblastos para su análisis. Las BMSC murinas se aíslan de huesos largos, se cultivan y se siembran en placas de 96 pocillos a una densidad de 25,000 células / pozo. El cultivo de estas células en medios específicos de osteoblastos se inicia cuando alcanzan el 80%-100% de confluencia para iniciar su diferenciación. Los ensayos se realizan en diferentes etapas de diferenciación. Las placas de los cartuchos se hidratan un día antes del ensayo. El día del ensayo, se inyectan diferentes inhibidores en los puertos de los cartuchos del sensor según los requisitos del ensayo, y se agrega un búfer de calibración a la placa de calibración de 96 pocillos. Después de la calibración, se realiza el ensayo de flujo metabólico celular en tiempo real, seguido de la obtención de imágenes de la microplaca de cultivo celular utilizando el generador de imágenes de microplacas para normalizar los datos del analizador de flujo metabólico celular en tiempo real con el recuento de células. Haga clic aquí para ver una versión más grande de esta figura.
