La formation d’os par l’ostéoblaste s’accompagne d’une destruction ou d’une résorption coordonnée des os par les ostéoclastes. L’équilibre entre la formation osseuse ostéoblastique et la résorption des ostéoclastes est un processus couplé décrivant le renouvellement osseux ou le remodelage, ce qui est essentiel pour l’homéostasie squelettique. Le dysfonctionnement ostéoblastique entraîne une altération de la formation osseuse et entraîne diverses maladies, y compris l’ostéoporose 1,2,3. La différenciation ex vivo/in vitro des cellules souches stromales de la moelle osseuse (BMSC) en précurseurs d’ostéoblastes et en ostéoblastes matures entraîne la formation et le dépôt de la matrice osseuse minéralisée dans le récipient de culture au fil du temps 4,5,6. Cette formation osseuse par l’ostéoblaste nécessite une quantité importante d’énergie cellulaire. Plus précisément, il a été démontré que la synthèse et la sécrétion de collagène dépendent fortement des rapports cellulaires ATP: ADP, et vraisemblablement, le trafic et la sécrétion de vésicules minéralisées nécessitent de l’ATP supplémentaire 7,8,9,10,11. De nombreux chercheurs ont démontré que le processus d’ostéoblastogenèse et de fonction ostéoblastique nécessite un apport adéquat d’énergie pour répondre à la demande métabolique de la formation osseuse 12,13,14,15,16. Par conséquent, l’objectif de cette méthode est de caractériser l’état bioénergétique des cellules stromales murines primaires tout au long de la différenciation ostéoblastique à l’aide de l’analyseur de flux métabolique cellulaire en temps réel. Ces techniques aident à développer une meilleure compréhension de l’homéostasie squelettique, ce qui pourrait finalement conduire au développement de nouvelles options thérapeutiques capables d’améliorer les troubles squelettiques.
L’analyseur de flux métabolique cellulaire en temps réel peut être utilisé pour mesurer le taux de consommation d’oxygène (OCR) et le taux d’acidification extracellulaire (ECAR) des ostéoblastes vivants, ce qui correspond à la production mitochondriale et glycolytique d’ATP, respectivement. Le fait qu’un ion H+ par lactate soit libéré lors de la glycolyse lors de la conversion du glucose en lactate, ce qui modifie le pH du milieu reflété dans les valeurs ECAR est fondamental pour cette méthodologie. Inversement, au cours du cycle TCA (acide tricarboxylique), la phosphorylation oxydative via les mitochondries produit du CO2 en utilisant ou en consommant de l’oxygène, et donc la surveillance de l’OCR reflète ce processus métabolique. L’analyseur mesure à la fois l’OCR et l’ECAR dans le microenvironnement extracellulaire simultanément et en temps réel, ce qui permet un potentiel énorme lors de l’étude de la bioénergétique cellulaire 6,17. De plus, l’exécution de ces tests est relativement simple et facilement personnalisable en fonction de l’objectif expérimental. Des techniques similaires ont été utilisées pour mieux comprendre la régulation métabolique des lymphocytes T du système immunitaire18,19, l’initiation et la progression du cancer20, ainsi que de multiples autres types de cellules contribuant aux syndromes métaboliques21,22.
Les avantages de l’analyseur de flux métabolique en temps réel par rapport aux techniques alternatives comprennent (1) la capacité de mesurer la bioénergétique cellulaire de cellules vivantes en temps réel, (2) la capacité d’effectuer des essais avec un nombre relativement faible de cellules (nécessite aussi peu que 5 000 cellules), (3) des ports d’injection pour manipuler en parallèle plusieurs traitements dans un système à haut débit de 96 puits, (4) l’utilisation d’un imageur de cellules automatisé sans étiquette radioactive pour la normalisation18, 23,24. Les méthodes suivantes visent à fournir une description généralisée mais détaillée de la surveillance de la bioénergétique cellulaire dans les BMSC murins tout au long de la différenciation des ostéoblastes à l’aide de l’analyseur. Il comprendra des tests effectués régulièrement; cependant, comme pour de nombreuses techniques et méthodes, il est fortement encouragé que les laboratoires individuels déterminent des détails spécifiques pour leurs expériences.
Sélection de tests et différents types de tests disponibles: Une grande variété de kits de dosage et de réactifs sont disponibles pour étudier la bioénergétique des cellules tout en assurant la fiabilité et la cohérence des résultats expérimentaux. En outre, le logiciel de bureau propose également des modèles de test qui peuvent être facilement personnalisés. Le test peut être défini en fonction des besoins de l’utilisateur pour mesurer différents paramètres métaboliques. Ces essais peuvent être modifiés de différentes manières en fonction de l’objectif expérimental et/ou de la question scientifique. Par exemple, avec quatre ports d’injection, plusieurs composés peuvent être injectés dans le milieu d’essai pour analyser la réponse cellulaire spécifique à chaque voie métabolique.
Test de phénotype d’énergie cellulaire: Ce test mesure le phénotype métabolique et le potentiel métabolique des cellules vivantes. Ce test est également recommandé comme première étape pour avoir une idée généralisée du métabolisme spécifique à la voie. Un mélange d’oligomycine A-un inhibiteur de l’ATP synthase et de cyanure de carbonyle 4-(trifluorométhoxy)phénylhydrazone (FCCP)-un agent de découplage mitochondrial est injecté pour comprendre le potentiel énergétique cellulaire. L’injection d’oligomycine A inhibe la synthèse de l’ATP, entraînant une augmentation du taux de glycolyse (ECAR) pour permettre aux cellules de répondre à leurs besoins énergétiques; d’autre part, l’injection de FCCP entraîne une OCR plus élevée en raison de la dépolarisation de la membrane mitochondriale. Essentiellement, ce test décrit la respiration métabolique basale et, après les doubles injections, poussées ou stress, la réponse métabolique. Sur la base de ces paramètres, le logiciel trace ensuite l’OCR et l’ECAR des cellules en classant les cellules en état aérobie, quiescent, glycolytique ou énergétique au fil du temps25,26.
Test du taux de production atp en temps réel : Cela mesure la production cellulaire d’ATP simultanément à partir de la glycolyse et de la respiration mitochondriale. Ce test mesure quantitativement les changements métaboliques des deux voies énergétiques et fournit des données sur les taux de production d’ATP mitochondrial et glycolytique au fil du temps. Le test obtient des données OCR et ECAR basales suivies du calcul du taux de production mitochondrial d’ATP par injection d’oligomycine A et du taux de production d’ATP glycolytique par injection de mélange roténone + antimycine A (inhibition totale de la fonction mitochondriale), entraînant une acidification mitochondriale17,27.
Test de stress des mitochondries cellulaires (ou test d’effort mito cellulaire): Cela mesure la fonction mitochondriale par la respiration liée à l’ATP, quantifie la bioénergétique cellulaire, identifie le dysfonctionnement mitochondrial et mesure la réponse des cellules au stress. Divers paramètres, y compris la capacité respiratoire basale et de rechange, la respiration liée à l’ATP, la respiration maximale et la consommation d’oxygène non mitochondriale, peuvent être obtenus en un seul test. Ce test implique des injections séquentielles d’oligomycine A, fccP (agent de découplage mitochondrial), un mélange d’inhibiteurs de roténone / antimycine A pour analyser efficacement l’effet de ceux-ci sur la fonction mitochondriale28.
Flexibilité mito fuel flex test: Cela mesure le taux de respiration mitochondriale par l’oxydation des trois combustibles mitochondriaux primaires par la présence et l’absence de leurs inhibiteurs. L’inhibition séquentielle du glucose, de la glutamine et des acides gras aide à mesurer la dépendance, la capacité et la flexibilité des cellules et la dépendance des cellules dans diverses voies cellulaires pour répondre à la demande d’énergie. Lorsque les mitochondries ne peuvent pas répondre aux exigences de la voie d’intérêt bloquée en oxydant d’autres combustibles, les cellules entrent dans un état de dépendance. La capacité des cellules est calculée par inhibition des deux autres voies alternatives suivies de l’inhibition de la voie d’intérêt. La flexibilité des cellules aide à comprendre la capacité des mitochondries à compenser et à répondre aux besoins en carburant de la voie inhibée. Il est calculé en soustrayant la dépendance des cellules de la capacité des cellules. Trois inhibiteurs différents sont utilisés indépendamment ou sous forme de mélange de deux pour calculer efficacement les paramètres du test. L’acide 2-cyano-3-(1-phényl-1H-indol-3-yl)-2-propénoïque (UK5099) inhibe l’oxydation du glucose en bloquant le transporteur de pyruvate dans la glycolyse. Le sulfure d’éthyle bis-2-(5-phénylacétamido-1,3,4-thiadiazol-2-yl) (BPTES) inhibe la voie d’oxydation de la glutamine et l’étomoxir inhibe l’oxydation des acides gras à longue chaîne29.
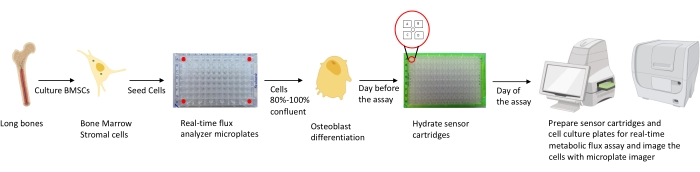
Figure 1 : Représentation schématique de la méthodologie de culture et de préparation des ostéoblastes pour analyse. Les BMSC murins sont isolés à partir d’os longs, cultivés et ensemencés dans des plaques de 96 puits à une densité de 25 000 cellules/puits. La culture de ces cellules dans des milieux spécifiques aux ostéoblastes commence lorsqu’elles atteignent une confluence de 80% à 100% pour commencer leur différenciation. Les tests sont effectués à différents stades de différenciation. Les plaques de cartouche sont hydratées un jour avant le test. Le jour du test, différents inhibiteurs sont injectés dans les orifices des cartouches de capteur en fonction des exigences du test, et un tampon d’étalonnage est ajouté à la plaque d’étalonnage de 96 puits. Après l’étalonnage, le test de flux métabolique cellulaire en temps réel est effectué, suivi de l’imagerie de la microplaque de culture cellulaire à l’aide de l’imageur de microplaques pour normaliser les données de l’analyseur de flux métabolique cellulaire en temps réel avec le nombre de cellules. Veuillez cliquer ici pour voir une version agrandie de cette figure.
