Barreiras apertadas separam o sistema nervoso central (SNC) da periferia, incluindo a barreira hematoencefálica (BHE) e a barreira sangue-líquido cefalorraquidiano (LCR). Essas barreiras protegem o SNC contra insultos externos e garantem um microambiente equilibrado e controlado 1,2,3. Embora a BHE tenha sido extensivamente estudada ao longo do tempo, a barreira sangue-LCR localizada no plexo coroide (CP) só ganhou crescente interesse de pesquisa durante a última década. Esta última barreira pode ser encontrada nos quatro ventrículos cerebrais (Figura 1A, B) e é caracterizada por uma única camada de células epiteliais do plexo coroide (ECP) ao redor de um estroma central, capilares vazados, fibroblastos e uma população de células linfoides e mieloides (Figura 1C)4,5,6. As células CPE estão firmemente interconectadas por tight junctions, evitando assim o vazamento dos capilares sanguíneos fenestrados subjacentes para o LCR e o cérebro. Além disso, o transporte através das células CPE é regulado por uma série de sistemas de transporte para dentro e para fora que gerenciam o influxo de compostos benéficos (por exemplo, nutrientes e hormônios) do sangue para o LCR e o efluxo de moléculas nocivas (por exemplo, resíduos metabólicos, excesso de neurotransmissores) na outra direção 1,6. Para poderem exercer sua função de transporte ativo, as células CPE contêm numerosas mitocôndrias em seu citoplasma7. Além disso, o PC é a principal fonte de LCR e atua como gatekeeper do cérebro pela presença de células inflamatóriasresidentes1. Devido à sua localização única entre o sangue e o cérebro, o PC também está perfeitamente posicionado para realizar vigilância imunológica8.
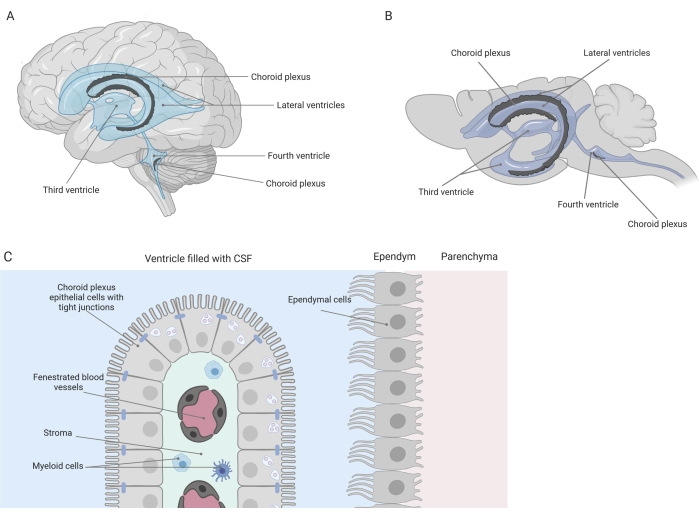
Figura 1: Visão geral esquemática da localização e composição do plexo coroide (PC). (A,B) O tecido CP é encontrado nos dois ventrículos laterais, o terceiro e o quarto ventrículos de (A) cérebro humano e (B) de camundongo. (C) O tecido do PC consiste de uma única camada de células do epitélio cuboidal do CP (EPC) firmemente conectadas ao redor de capilares fenestrados, tecido conjuntivo frouxo e células linfoides e mieloides, e forma a barreira hemato-liquórica (adaptada e modificada da referência23). Figura criada com Biorender.com. Clique aqui para ver uma versão maior desta figura.
Na última década, evidências crescentes, incluindo vários relatos de nosso grupo de pesquisa, têm revelado que a PC desempenha um papel central na saúde ena doença9,10,11,12,13,14,15,16,17,18 . Por exemplo, sabe-se que a barreira sangue-LCR do envelhecimento apresenta alterações morfológicas, entre outras, nos núcleos, microvilosidades e membrana basal 1,19. Além disso, no contexto da doença de Alzheimer, a integridade global da barreira está comprometida e todas essas mudanças relacionadas à idade parecem ser ainda mais pronunciadas 1,8,20. Além das alterações morfológicas, o transcriptoma, proteoma e secretoma da PC são alterados durante a doença 12,21,22,23. Assim, o conhecimento avançado da PC é essencial para melhor compreender seu papel nas doenças neurológicas e potencialmente desenvolver novas estratégias terapêuticas.
Um método eficiente para a microdissecção precisa do PC para fora dos ventrículos cerebrais é o primeiro passo inestimável para permitir a investigação adequada desta minúscula estrutura cerebral. Devido à sua natureza altamente vascularizada (Figura 2B), o CP flutuando dentro das cavidades ventriculares do encéfalo pode ser identificado por meio de um microscópio binocular. No entanto, a perfusão transcárdica é frequentemente necessária para análise a jusante, dificultando a identificação e o isolamento adequados do tecido do PC (Figura 2C). Se as etapas adicionais de processamento permitirem (por exemplo, no caso de análise de RNA e proteínas), o CP pode ser visualizado via perfusão transcárdica com azul de bromofenol (Figura 2A). Várias publicações já descrevem o isolamento do PC do cérebro de ratos24 e filhotes de camundongos25. Aqui, uma técnica de isolamento por microdissecção é descrita para isolar o PC de camundongos adultos. É importante ressaltar que essa técnica de isolamento preserva a viabilidade, a função e a estrutura das células dentro do PC. O isolamento do CP flutuando no quarto e nos ventrículos laterais é descrito aqui. Em suma, os camundongos são anestesiados terminalmente e, se necessário, perfundidos transcardialmente. No entanto, deve-se notar que a perfusão pode danificar a estrutura das células dentro do PC. Consequentemente, se a amostra for analisada usando microscopia eletrônica de transmissão (MET), microscopia eletrônica de varredura de face serial block (SBF-SEM) ou MEV por feixe de íons focalizado (MEV-FIB), a perfusão não deve ser realizada. Em seguida, todo o cérebro é isolado, e fórceps são usados para hemissectar sagitalmente o cérebro. A partir daí, os CPs que flutuam nos ventrículos laterais podem ser identificados e dissecados, enquanto os CP do quarto ventrículo podem ser isolados do lado cerebelar do cérebro.
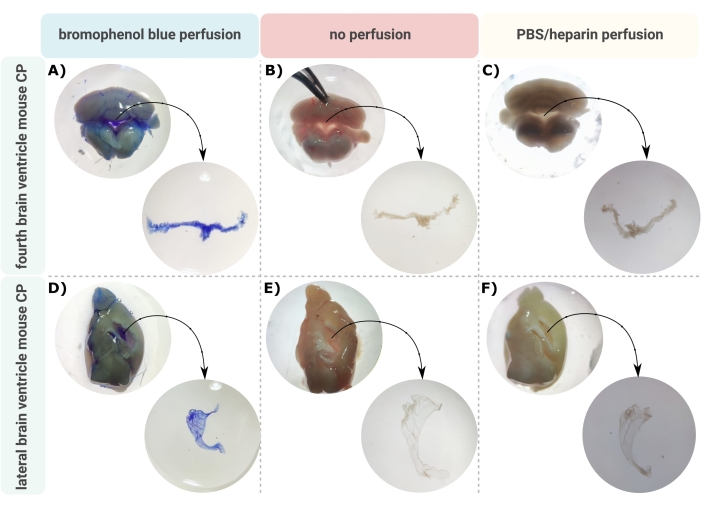
Figura 2: Visualização do plexo coroide (CP) do quarto e (D-F) ventrículo lateral (CP) após (A,D) perfusão com azul de bromofenol, (B,E) sem perfusão e (C,F) perfusão com PBS/heparina. As imagens são obtidas com estereomicroscópio (aumento de 8x-32x). Clique aqui para ver uma versão maior desta figura.
Uma vez que o PC é adequadamente dissecado para fora dos ventrículos cerebrais, todo um repertório de técnicas pode ser aplicado para obter maior compreensão sobre a função dessa estrutura. Por exemplo, citometria de fluxo ou sequenciamento de RNA de célula única podem ser realizados para quantificar e analisar fenotipicamente as células inflamatórias infiltrantes sob determinadas condições de doença26,27. Além da composição celular, a composição molecular do CP pode ser analisada para avaliar a presença de citocinas e quimiocinas via ensaio imunoenzimático (ELISA), immunoblot, ou através da análise simultânea de múltiplas citocinas utilizando o arranjo de esferas de citocinas28. Além disso, análises de transcriptoma, vascular, histologia de células imunes e secretoma podem ser realizadas nos explantes de CPmicrodissecados29. Aqui, a microscopia eletrônica de varredura (MEV) em CP de montagem inteira é usada para obter uma visão geral da estrutura do CP. O MEV usa um feixe de elétrons focalizados para varrer a superfície e criar uma imagem da topografia e composição da superfície. Como o comprimento de onda dos elétrons é muito menor do que o da luz, a resolução do MEV está na faixa nanométrica e é superior à de um microscópio de luz. Consequentemente, estudos morfológicos em nível subcelular podem ser realizados via MEV. Resumidamente, o CP dissecado é imediatamente transferido para um fixador contendo glutaraldeído para uma fixação noturna, seguida de osmicação e coloração com acetato de uranila. As amostras são então tratadas com coloração de aspartato de chumbo, desidratadas e, finalmente, incluídas para aquisição de imagens.
Assim, este protocolo facilita o isolamento eficiente do PC dos ventrículos cerebrais de camundongos, o que pode ser posteriormente analisado usando uma variedade de técnicas a jusante para investigar sua estrutura e função.
