We previously documented biliary defects in Jag1+/– animals, a mouse model of ALGS8. To determine the BD to PV ratio, we sectioned P30 mouse livers and co-stained them for CK8 and CK19 (wsCK) along with the vascular marker αSMA. We then imaged all the PVs in each of the liver lobes. As shown in Figure 2A, we defined PVs as αSMA-stained vessels that have adjacent wsCK staining (arrowheads). The αSMA-stained structures without wsCK were central veins and should not be included in the analysis (arrow).
Once PVs were identified, we identified patent BDs by their characteristic shape. As shown in Figure 3, patent ducts have a clearly definable lumen that is surrounded by wsCK+ cholangiocytes. The ducts are usually separated from nearby ducts or cholangiocytes by mesenchyme (arrowhead). wsCK+ cells that do not have a definable lumen, are attached to adjacent cells, or appear in isolation, are not counted toward the total number of BDs (arrows). Figure 3A shows a wild-type liver section with a PV which is associated with a fully patent duct along with several unincorporated cells. Figure 3B is a representative liver section from a P30 Jag1+/– animal. No patent BDs are present around the three PVs in this section. All wsCK+ cells are unincorporated and therefore should not be counted. This image highlights the importance of careful BD counting, as presence or absence of wsCK+ cells does not differentiate the Jag1+/– and wild-type livers.
As demonstrated in Figure 1D, analysis of the BD to PV ratio involves counting every PV in the liver section along with the total number of patent BDs present around each PV. While analyzing the Jag1+/– livers, we noticed that different lobes are not necessarily affected to the same extent in these animals (unpublished data). Therefore, we usually count PVs across the left, medial, right and caudate lobes to ensure complete liver coverage. Following tabulation of total PVs and BDs, the ratio is calculated for the whole section.
In Figure 4, we showed the analysis of the BD to PV ratios for 3 wild-type and 3 Jag1+/– animals. This graph shows how the two genotypes can be readily distinguished based on BD counts. Additionally, this method provides a quantitative measure for analysis of the degree of rescue of the Jag1+/– phenotype by genetic manipulations, as reported previously8.
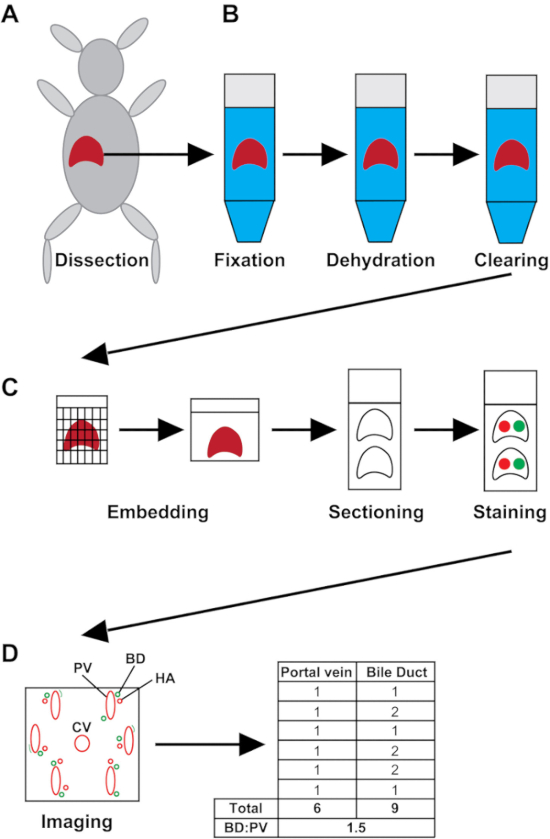
Figure 1: Schematic of the experimental process. (A) The liver is harvested whole from the mouse. (B) Liver samples are fixed for 48 h, dehydrated and cleared. (C) The liver tissue is embedded in paraffin. Sections are made and placed on charged slides and stained for wsCK and αSMA. (D) Slides are imaged, and the number of PVs and BDs are recorded. The BD to PV ratio (BD:PV) is calculated for the entire liver section. HA = hepatic artery; CV = central vein. Please click here to view a larger version of this figure.
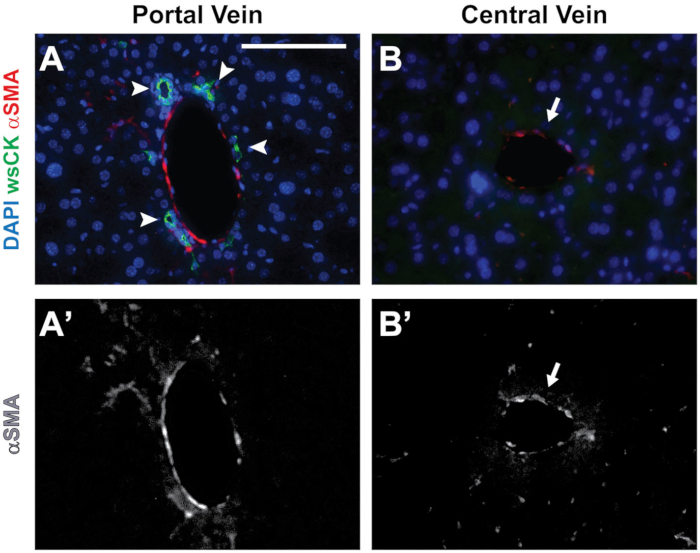
Figure 2: Distinguishing central veins from PVs. (A) A PV is identified by the presence of αSMA staining and surrounding wsCK+ cholangiocytes (arrowheads). (B) Central veins are identified by the presence of αSMA staining with no wsCK+ cholangiocytes present around the structure (arrow). (A’ and B’) Grayscale images showing the αSMA channels from A and B, respectively. Scale bar = 100 µm and applies to all panels. Please click here to view a larger version of this figure.
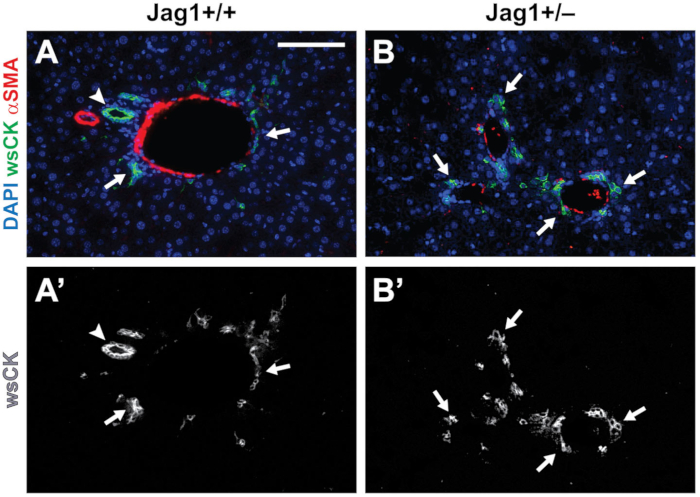
Figure 3: Identification of patent BDs. (A-A’) In P30 wild-type livers, we count round to ellipsoid structures with a discernable lumen surrounded by wsCK+ cholangiocytes as a patent BD (arrowhead). Cholangiocytes which are not surrounding a lumen are considered unincorporated and are not counted (arrows). (B-B’) In Jag1+/– livers, cholangiocytes are still present (arrows). However, most are not incorporated into patent BDs. Scale bar = 100 µm and applies to all panels. Please click here to view a larger version of this figure.
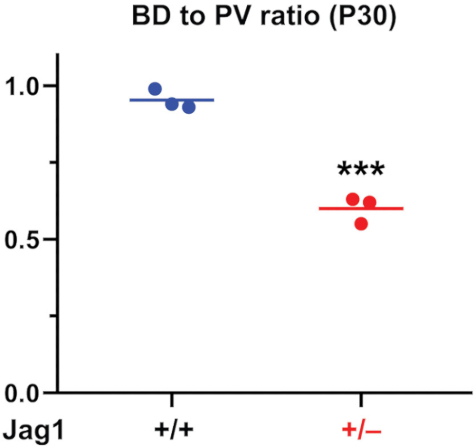
Figure 4: The BD to PV Graph from P30 mouse livers. BDs and PVs are quantified and the BD to PV ratio is generated. As reported previously8, Jag1+/– animals have a characteristic and significant decrease in BD to PV ratio compared to wild-type animals. For statistical analysis, two-way t-test was performed. Horizontal lines show means. *** P<0.001. Please click here to view a larger version of this figure.
