This protocol presents procedures for generating a homogenous culture of spheroids from primary tumor cells, quantitatively evaluating drug efficacy on spheroid culture (MTT assay), and determining the effect of study drugs on spheroid morphology. Data from the representative experiments in spheroids generated from colon and breast cancer cell cultures are presented. Similar experiments were performed using other tumor types, including cholangiocarcinoma, gastric, lung, and pancreatic cancer (data not shown). All the experiments presented herein were performed in triplicate.
Figure 2 shows the spheroids that were generated from the primary colon cancer cell culture. As seen in Figure 2, the number of spheroids generated depends on the number of cells initially seeded in each well. The growth of the spheroids to over 100 μm in diameter took 10-14 days. The origin of the tumor cells (e.g., different patients and different origins) determined the growth rate. Seeding the wells with more cells did not shorten the time required for spheroid generation but rather increased the number of spheroids formed. Notably, upon prolonged culture of the colon cancer spheroids, they started attaching to each other and formed clusters of spheroids in grape-like structures (Figure 3), which prevented a homogenous culture and, thus, prohibited the use of the spheroids in the MTT assays.
Figure 4 presents the effect of three treatments (10 μM palbociclib, 10 μM sunitinib, and their combination at 10 μM each) on the viability of spheroids derived from two primary cancers. In this case, a PDX model was established first, and the tumor cells used for the spheroid analysis were derived from the PDX model9. The first PDX model was established using a colon cancer sample from a 50 year old male patient, and the second using a breast cancer sample from a 62 year old female. As demonstrated in Figure 4A,B, after 3 days of treatment, the combination of palbociclib plus sunitinib led to a significant reduction in viability as measured by the MTT assay. As demonstrated in Figure 4C,D, the morphological changes occurring with treatment were very clear. On Day 0, all the spheroids were intact. In contrast, on Day 3, the spheroids treated with the control (DMSO) were still intact, whereas the spheroids treated with the combination were disassembled, and their morphology was "open", with cells detaching from the solid structure, suggesting the destruction of the spheroid structure.
Figure 5 presents the follow-up of the spheroids over time. These spheroids, generated from breast cancer cells derived from a 44 year old female patient, were treated with one of two combinations (trastuzumab [10 μg/mL] plus vinorelbine [1 μg/mL], or 5-fluorouracil [200 μM] plus cisplatin [300 μM]). As shown in Figure 5A, the size of the spheroids treated with 5-fluorouracil plus cisplatin was reduced by Day 3, and the spheroids were completely destroyed by Day 7. In contrast, the treatment with trastuzumab plus vinorelbine had only a minor effect on the morphology of the spheroids (e.g., some level of an "open" structure), but the effect was not significant. Figure 5B presents the average change in the diameter of the spheroids relative to Day 0 (five spheroids were followed in each treatment group).
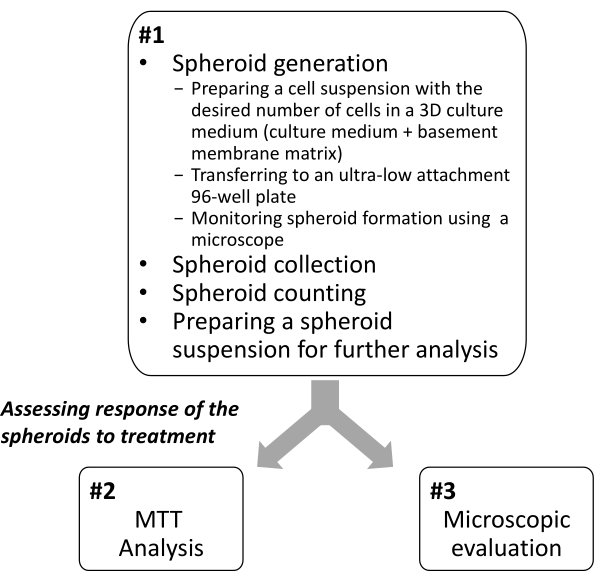
Figure 1: Overview of the protocol for establishing 3D spheroids from patient-derived tumor samples and evaluating their sensitivity to drugs. Please click here to view a larger version of this figure.
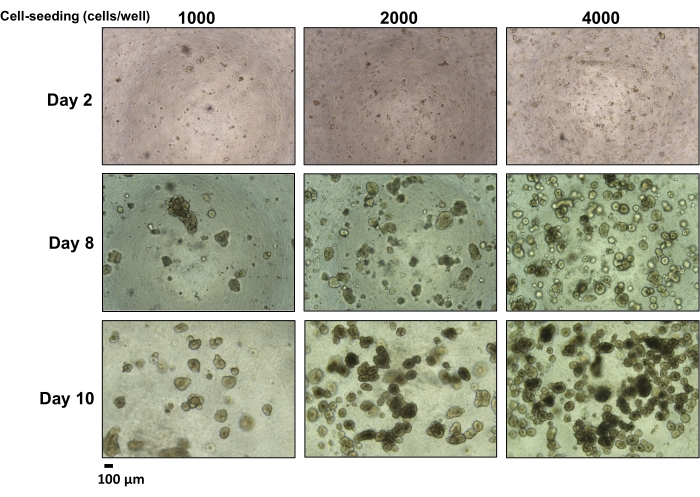
Figure 2: Formation of spheroids from a primary colon cancer cell culture over time by the number of initially seeded cells. Different numbers of cells were seeded in "3D culture medium" in an ultra-low attachment 96-well plate and observed under the microscope (4x magnification). Scale bar = 100 µm. Please click here to view a larger version of this figure.
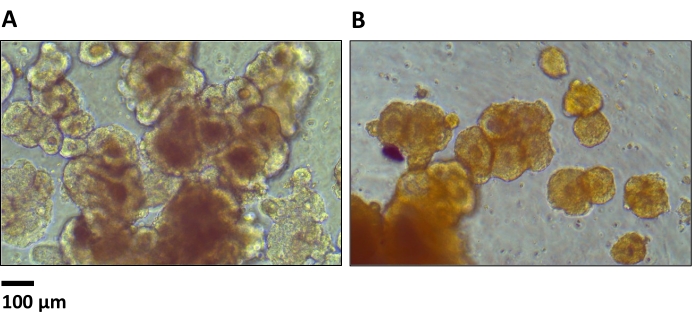
Figure 3: Spheroids from primary colon cancer cells (with initial cell seeding of 2,000 per well) after 12 days in culture. The two examples (A,B) show clusters created by the attachment of spheroids to each other (10x magnification). Scale bar = 100 µm. Please click here to view a larger version of this figure.
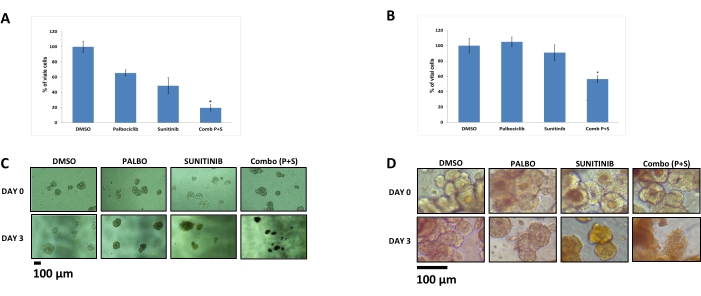
Figure 4: The effects of palbociclib (10 μM), sunitinib (10 μM), and their combination (10 μM each) on spheroids from primary tumor cells, including colon and breast cancer (PDX-derived). An MTT assay was conducted on spheroids derived from (A) colon and (B) breast cancer cells. The MTT signals were normalized to values from DMSO-treated cells. The values represent the means from four to eight replicates. The error bars represent SEM. *p < 0.05 versus a single agent (t-test). The effects of the various treatments on cell growth were also evaluated microscopically at Day 0 and after 3 days of treatment of the spheroids derived from (C) colon and (D) breast cancer cells (10x magnification). Scale bar = 100 µm. The figure is adapted from Moskovits et al.9. Please click here to view a larger version of this figure.
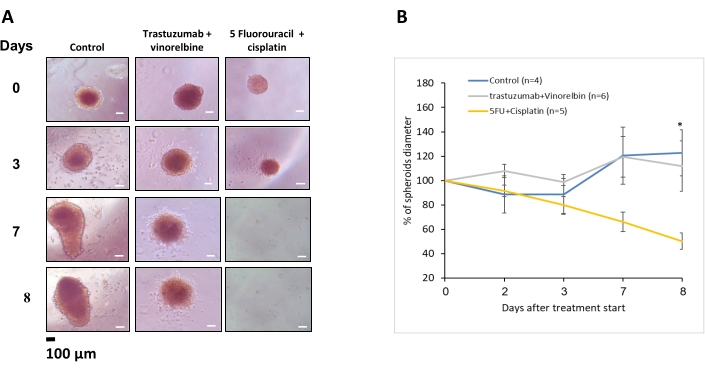
Figure 5: The effects of trastuzumab (10 μg/mL) plus vinorelbine (1 μg/mL) and 5-fluorouracil (200 μM) plus cisplatin (300 μM) on spheroids derived from breast cancer over time. (A) Each well included one spheroid and was monitored over time under the microscope (10x magnification). Scale bar = 100 µm. (B) The change in the diameter of the spheroids (relative to Day 0) by the duration of treatment. *p = 0.05 for 5-fluorouracil plus cisplatin versus controls (t-test). Each treatment group included four to six wells, with one spheroid in each well. The average change is presented. The error bars represent SEM. Please click here to view a larger version of this figure.
