L’OS est un type de tissu conjonctif composé de deux parties : organique (les cellules et les fibres de collagène) et minéraux (composés de calcium et de phosphate). Les principaux éléments minéraux dans les os sont apatites1. Différents types de cellules capables de minéralisation osseuse (ostéoblastes), dans les dents (odontoblastes) et dans le cartilage (chondrocytes) réglementent les étapes initiales de la minéralisation en produisant des protéines de la matrice extracellulaire (ECM) et en relâchant la matrice vésicules (MVs) (Figure 1). MVs sont 100-300 nm de diamètre vésicules qui s’accumulent de calcium et phosphate facilitant la nucléation d’apatite et par la suite se lient au collagène2,3. Puis, MVs se désagrègent pour relâcher les apatites dans le milieu extracellulaire. Les apatites continuent de croître en contact avec les fibres de collagène et de former la matrice osseuse. La minéralisation est soutenue par l’apport constant de Pj’ai et Ca2 + dans le milieu extracellulaire. Certaines données publiées récemment soutiennent notre modèle4,5. Tissus mous minéraliser pas dans des conditions physiologiques. Cependant, les calcifications ectopiques peuvent survenir dans des conditions pathologiques telles que la calcification vasculaire3. Les cellules vasculaires qui acquièrent le phénotype ostéoblastique peuvent produire MVs qui induisent la nucléation des apatites et initient la minéralisation dans les couches médianes et intima de la paroi des vaisseaux sanguins. Depuis les calcifications ectopiques ressemblent à des normales endochondrale minéralisation3, comprendre les mécanismes moléculaires de la minéralisation des cellules osseuses et chondrocytes devrait fournir quelques indices sur la calcification des tissus mous qui sont ectopique formé.
Le développement des tissus osseux est régi par diverses enzymes, des facteurs de croissance et des promoteurs ou des inhibiteurs de minéralisation. L’action antagoniste du tissu non-spécifique à la phosphatase alcaline (SERVT) (Figure 1) et ectonucleotide pyrophosphatase/phosphodiestérase j’ai (NPP1), ainsi que de l’ankyrin (ANK), contrôle concentration pyrophosphate inorganique (PPj’ai) 6. PPj’ai, un puissant inhibiteur de la formation de HA, est hydrolysé par SERVT ; NPP1 hydrolyse triphosphates de nucléotides pour former PPj’ai tandis que ANK PP j’exporte de la cellule à l’ECM. Le ratio de Pi/PPi peut réglementer apatite formation7,8 , avec des conséquences pathologiques possibles9.
La membrane de MV est enrichie en protéines de transport des ions qui facilitent la précipitation initiale du calcium et du phosphate à l’intérieur de la VM au cours du processus de nucléation (Figure 1). Le transporteur de phosphate 1 (PiT) permet d’intégrer Pj’ai généré dans l’espace perivesicular dans le MVs10,11. ANNEXINES peuvent être impliqués dans la liaison et transport de Ca2 + et dans le processus biophysique qui initie la minéralisation dans le MV lumen12,13. Nous sommes favorables à l’hypothèse, suggéré plus tôt, pour la minéralisation dans les vésicules intracytoplasmiques de nucléation interne d’apatite dans le MV avant sa propagation dans l’ECM14,15. In vitro de modélisation a confirmé l’induction de Ca2 +/ pje complexes formation en protéoliposomes fait du PS et AnxA516. Cela peut indiquer que l’accumulation de Ca2 +, Pi, AnxA5 et PS complexes en radeaux lipidiques des microvillosités-comme membranesrepresent le noyau de la nucléation (NC) d’apatite dans Mvs ANNEXINES et SERVT possèdent également liant le collagène capacités qui peuvent être utiles en plaçant MVs le long des fibres de collagène et en stimulant la propagation de la minéralisation de la MEC. Fetuin A et ostéopontine (OPN)17, sont connus comme inhibiteurs de la formation d’apatite qui peuvent ralentir la propagation de la minéralisation à l’échafaud collagénique. Nucléation et propagation sont des événements distincts, le premier qui précède ce dernier, et les deux peuvent être pertinents pour le processus de minéralisation pathologique.
Pour découvrir comment la chimie des complexes de phosphate de calcium peut changer minéralisation physiologique et calcification ectopique, il est nécessaire d’identifier les minéraux produits de cellules. Apatites sont un groupe de calcium et de phosphate contenant des minéraux avec le cristal générales unité cellulaire formule Ca10(PO4)6X2, où X = Cl, F, OH. Ils sont classés comme suit18: fluorapatite (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 et hydroxyapatite (HA) Ca10(PO4 )6(OH)2.
Le choix des lignées de cellules ostéoblastes à induire la formation de minéraux est crucial, puisque chaque lignée cellulaire montre un profil distinct de minéralisation. Dans ce rapport, nous avons comparé la nucléation des minéraux par deux modèles de cellule humaine de minéralisation : cellules ostéoblastique hFOB 1.19 et ostéosarcome HRA-2. Cellules dérivées d’ostéosarcome sont couramment utilisés comme modèles ostéoblastique et la Saos-2 cellules ont conservé le plus mature de caractère ostéoblastique19 tandis que les cellules indifférenciées hFOB foetale humaine sont largement utilisés comme modèle pour la normale ostéoblastique 20de différenciation. Leurs profils de minéralisation ont été analysés par différentes méthodes : alizarine rouge-S (AR-S) coloration, ultraviolette visualisation de lumière (UV), imagerie de microscopie électronique (met) de transmission, énergie dispersive x-ray microanalysis (EDX) quantification et ion cartographie. L’avantage de TEM-EDX sur les autres techniques utilisées dans les études précédentes est qu’il donne des résultats quantitatifs et qualitatifs de remplacement ion en apatite cristaux4,5,21. L’objectif global de l’utilisation de TEM-EDX était de trouver une méthode simple pour l’imagerie et la quantification de la répartition des ions Ca, F et Cl dans divers minéraux de différents types de cellules au cours des étapes distinctes du processus de minéralisation. Cette méthode a été utilisée avec succès, par exemple, pour surveiller l’interaction des nanoparticules de zinc avec des produits chimiques coexistantes et leurs effets combinés sur les organismes aquatiques,22. Dans une autre étude, un PHOTOCATALYSEUR cuivre sur matériaux titane en solution aqueuse a été largement caractérisée par spectrométrie d’émission optique de plasma à couplage inductif (ICP-OES), N2 physisorption (BET), XRD, DRS UV-vis, FT-IR, Raman spectroscopie, TEM-EDX et photoélectrochimiques mesures23. Notre but était de comparer l’origine et les propriétés de vésicules et de minéraux dans les deux lignées cellulaires pour comprendre le mécanisme qui contrôle la minéralisation durant la différenciation osseuse.
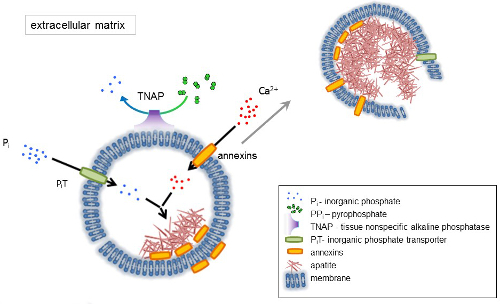
Figure 1 . Schéma des étapes initiales de la minéralisation dans les cellules osseuses impliquant la synthèse des protéines de la matrice extracellulaire (ECM) et la libération des vésicules (MVs) de la matrice de la membrane. MVs accumulent calcium grâce à l’action des protéines liant le calcium, d’ANNEXINES et de phosphate, par l’action d’un transporteur de phosphate inorganique (PiT) suivi de l’activité du tissu non spécifiques à la phosphatase alcaline (SERVT), qui dephosphorylates PPj’ai Pje, facilitant ainsi la nucléation d’apatite. Ensuite, MVs se désintègrent et libérer des apatites dans le milieu extracellulaire. La minéralisation est soutenue par l’apport constant de Pj’ai et Ca2 + dans le milieu extracellulaire4,5. S’il vous plaît cliquez ici pour visionner une version agrandie de cette figure.
