Osso é um tipo de tecido conjuntivo composto de duas partes: orgânico (células e fibras colágenas) e mineral (compostos de cálcio e fosfato). Os principais componentes minerais nos ossos são apatites1. Diferentes tipos de células competentes de mineralização no osso (osteoblastos), dentes (odontoblastos) e cartilagem (condrócitos) regulam os passos iniciais da mineralização através da produção de proteínas da matriz extracelular (ECM) e liberando a matriz vesículas (MVs) (Figura 1). MVs são vesículas de diâmetro de nm de 100-300 que acumulam cálcio e fosfato, facilitando a nucleação de apatita e posteriormente vincular a colágeno2,3. Em seguida, MVs desintegrar-se para liberar apatites para o meio extracelular. Os apatites continuam a crescer em contato com as fibras de colágeno e formam a matriz óssea. A mineralização é sustentada pela oferta constante de Peu e Ca2 + no meio extracelular. Alguns dados publicados recentemente apoiar nosso modelo4,5. Tecidos moles não mineralize sob condições fisiológicas. No entanto, calcificação ectópica pode ocorrer sob condições patológicas como calcificação vascular3. Células vasculares que adquirem o fenótipo de osteoblastos podem produzir MVs que induzem a nucleação de apatites e iniciar a mineralização nas camadas da parede medial e da íntima dos vasos sanguíneos. Desde calcificação ectópica se assemelham a endocondral normal mineralização3, compreender os mecanismos moleculares da mineralização das células ósseas e condrócitos devem fornecer algumas pistas sobre gravidez ectópica calcificação dos tecidos moles que são formada.
O desenvolvimento dos tecidos esqueléticos é regulado por diversas enzimas, fatores de crescimento e promotores ou inibidores de mineralização. A ação antagonista de tecido-inespecíficos da fosfatase alcalina (TNAP) (Figura 1) e ectonucleotide pyrophosphatase/fosfodiesterase eu (NPP1), juntamente com ankyrin (ANK), controla a concentração de pirofosfato inorgânico (PPeu) 6. PPeu, um potente inibidor da formação de HA, é hidrolisado por TNAP; NPP1 hidrolisa trifosfatos de nucleotídeos para formar PPeu enquanto ANK exporta-PPeu da célula para o ECM. A proporção de Pi/PPi pode regular apatita formação7,8 , com possíveis consequências patológicas9.
A membrana de MV é rico em proteínas de transporte de íons que facilitam a precipitação inicial do cálcio e fosfato dentro os MVs durante o processo de nucleação (Figura 1). O transportador de fosfato 1 (PiT) ajuda a incorporar Peu gerados no espaço perivesicular para o MVs10,11. Anexinas podem estar envolvidas na ligação e transporte de Ca2 + e no processo de Biofísica que inicia a mineralização na MV lúmen12,13. Favorecemos a hipótese, sugerida anteriormente, para mineralização dentro de vesículas intracitoplasmática de nucleação interna de apatita dentro o MV antes de sua propagação na ECM14,15. Modelagem em vitro confirmou a indução de Ca2 +/ peu complexos formação em proteoliposomes feitas de PS e AnxA516. Isso pode indicar que a acumulação de Ca2 +, Peu, AnxA5 e PS complexos em balsas lipídicas de membranesrepresent microvilli-como o núcleo de nucleação (NC) de apatita dentro Mvs anexinas e TNAP também possuem ligação de colágeno capacidades que podem ser úteis na colocação de MVs ao longo de fibras de colagénio e, para estimular a propagação da mineralização em ECM. Fetuin A e osteopontin (OPN)17, são conhecidos como inibidores da formação de apatita que pode abrandar a propagação de mineralização no cadafalso colágenas. Nucleação e propagação são eventos distintos, o primeiro anterior a último, e ambos podem ser relevantes para o processo de mineralização patológica.
Para descobrir como a química de complexos de fosfato de cálcio pode alterar mineralização fisiológica e calcificação ectópica, é necessário identificar os minerais produzidos por células. Apatites são um grupo de cálcio e fosfato contendo minerais com cristal geral unidade célula fórmula Ca10(PO4)6X2, onde X = Cl, F, OH. Eles são classificados como segue18: fluorapatita (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 e hidroxiapatita (HA) Ca10(PO4 )6(OH)2.
A escolha de linhas de células osteoblast para induzir a formação de minerais é crucial, pois cada linha celular apresenta um perfil distinto de mineralização. Neste relatório, nós comparamos a nucleação de minerais por dois modelos de célula selecionada humana de mineralização: células osteoblásticas hFOB 1.19 e osteossarcoma Saos-2 células. Osteossarcoma-derivado de células são comumente usadas como modelos osteoblásticas e Saos-2 células têm preservado o caráter osteoblástica mais maduro19 enquanto células indiferenciadas hFOB fetal humano são amplamente utilizadas como um modelo para o normal osteoblástica diferenciação de20. Seus perfis de mineralização foram analisados por métodos diferentes: coloração vermelho de alizarina-S (AR-S), ultravioleta (UV) luz visualização, imagem de microscopia eletrônica (TEM) de transmissão, energia dispersiva raio-x (EDX) de microanálise quantificação da e íon mapeamento. A vantagem da temperatura-EDX sobre alternativas técnicas utilizadas em estudos anteriores é que dá resultados quantitativos e qualitativos de substituição iônica em cristais de apatita a4,5,21. O objetivo geral de usar TEM-EDX foi encontrar um método simples para geração de imagens e quantificação da distribuição dos íons Ca, F e Cl em vários minerais de diferentes tipos de células durante estágios distintos do processo de mineralização. Este método tem sido utilizado com sucesso, por exemplo, para monitorar a interação de nanopartículas de zinco com produtos químicos coexistentes e seus efeitos combinados em organismos aquáticos22. Em outro estudo, um photocatalyst cobre em materiais de titânio em solução aquosa foi extensivamente caracterizado por espectrometria de emissão óptica de plasma indutivo (ICP-OES), physisorption N2 (BET), XRD, DRS UV-vis, FT-IR, Raman espectroscopia, TEM-EDX e photoelectrochemical medidas23. Nosso objetivo foi comparar a origem e propriedades de vesículas e minerais em duas linhas de celular para entender o mecanismo que controla a mineralização durante a diferenciação óssea.
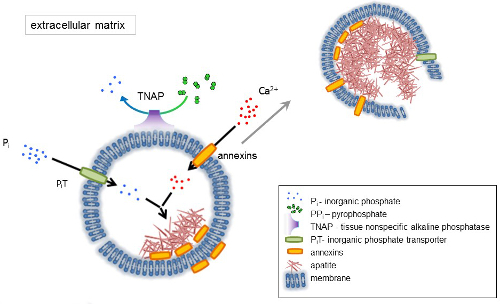
Figura 1 . Esquema das etapas iniciais de mineralização em células ósseas envolvendo a síntese de proteínas da matriz extracelular (ECM) e a liberação das vesículas de matriz (MVs) da membrana. MVs acumulam cálcio através da ação de proteínas de ligação de cálcio, anexinas e fosfato, através da ação do transportador de fosfato inorgânico (PiT) seguido pela actividade de tecido não-específica da fosfatase alcalina (TNAP), que dephosphorylates PPeu Peu, facilitando assim a nucleação de apatita. Em seguida, MVs desintegrar-se e liberar apatites para o meio extracelular. A mineralização é sustentada pela oferta constante de Peu e Ca2 + no meio extracelular4,5. Clique aqui para ver uma versão maior desta figura.
