Bot is een soort bindweefsel bestaat uit twee delen: organische (cellen en collageenvezels) en mineralen (calcium en fosfaat verbindingen). De belangrijkste minerale componenten in botten zijn apatites1. Verschillende soorten mineralisatie-bevoegde cellen in bot (botcellen), tanden (odontoblasts) en kraakbeen (chondrocyten) regelen de eerste stappen van mineralisatie door de productie van eiwitten van de extracellulaire matrix (ECM) en het vrijgeven van matrix blaasjes (MVs) (Figuur 1). MVs zijn 100-300 nm diameter blaasjes die zich ophopen van calcium en fosfaat te vergemakkelijken apatiet nucleatie en vervolgens binden aan collageen2,3. Vervolgens desintegreren MVs om apatites aan het extracellulaire medium vrij te geven. De apatites blijven groeien in het contact met de collageenvezels en vormen de bot-matrix. De mineralisatie is opgelopen door de constante aanvoer van Pik en Ca2 + in het extracellulaire medium. Sommige onlangs gepubliceerde gegevens ondersteunen onze model4,5. Zachte weefsels doen niet mineralize onder fysiologische omstandigheden. Ectopische verkalking kan zich echter voordoen onder pathologische omstandigheden zoals vasculaire verkalking3. Vasculaire cellen die het osteoblast fenotype verwerven kunnen produceren MVs dat induceren nucleatie van apatites en initiëren van mineralisatie in de mediale en intima lagen van de wand van de bloedvaten. Sinds ectopische verkalking lijken op normale endochondral mineralisatie3, inzicht in de moleculaire mechanismen van mineralisatie van ossaal cellen en chondrocyten dienen enkele aanwijzingen op ectopische verkalking van de zachte weefsels die gevormd.
De ontwikkeling van het skelet weefsels wordt geregeld door verschillende enzymen, groeifactoren, en initiatiefnemers of remmers van mineralisatie. De antagonistische actie van weefsel-nonspecific alkalisch fosfatase (TNAP) (Figuur 1) en ectonucleotide pyrophosphatase/fosfodiësterase ik (NPP1), samen met ankyrin (ANK), besturingselementen concentratie van de anorganische pyrofosfaat (PPik) 6. PPik, een krachtige remmer van HA formatie, is gehydrolyseerd door TNAP; NPP1 ontstabiel nucleotide trifosfaat tot PPik terwijl ANK uitvoer PPik uit de cel aan de ECM. De Pi/PPi verhouding kan reguleren apatiet vorming7,8 met mogelijke gevolgen van de pathologische9.
De MV membraan is verrijkt met ion vervoer eiwitten die het eerste neerslaan van calcium en fosfaat binnen de MVs tijdens de nucleatie (Figuur 1 vergemakkelijken). De fosfaat transporter 1 (PiT) helpt te nemen Pik gegenereerd in de perivesicular ruimte in de MVs10,11. Annexins kan worden betrokken in het bindende en vervoeren van Ca2 + en de biofysische proces waarmee mineralisatie in de MV lumen12,13. Wij keuren de hypothese, eerder voorgesteld voor mineralisatie binnen intracytoplasmatische blaasjes van interne nucleatie van apatiet binnen de MV voordat de vermeerdering in de ECM14,15. In vitro modellering bevestigd de inductie van Ca2 +/Pik complexen vorming in proteoliposomes gemaakt van PS en AnxA516. Dit kan erop duiden dat accumulatie van Ca2 +, Pi, AnxA5 en PS complexen in lipide vlotten van de microvilli-achtige membranesrepresent de nucleatie kern (NC) van apatiet binnen MVs. Annexins en TNAP bezitten ook collageen-bindende capaciteiten die nuttig zijn in het plaatsen van MVs langs collageenvezels en bij het stimuleren van de verspreiding van mineralisatie in de ECM kunnen zijn. Fetuin A en osteopontin (OPN)17, staan bekend als remmers van apatiet formatie die de verspreiding van mineralisatie op de collagene steiger kan vertragen. Nucleatie en vermeerdering zijn verschillende evenementen, de voormalige voorafgaand aan de laatste, en beide relevant kunnen zijn voor het proces van pathologische mineralisatie.
Om te ontdekken hoe de chemie van calciumfosfaat complexen fysiologische mineralisatie en ectopische verkalking kan veranderen, is het noodzakelijk om te identificeren van de mineralen geproduceerd door cellen. Apatites zijn een groep van calcium en fosfaat met mineralen met de algemene crystal eenheid cel Ca formule10(PO4)6X2, waarbij X = Cl, F, OH. Ze worden geclassificeerd als volgt18: fluorapatiet (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 en hydroxyapatiet (HA) Ca10(PO4 )6(OH)2.
De keuze van osteoblast cellijnen voor het opwekken van minerale vorming is van cruciaal belang, aangezien elke cellijn een duidelijk profiel van mineralisatie vertoont. In dit verslag, vergeleken we de nucleatie van mineralen door twee geselecteerde menselijke cel modellen van mineralisatie: osteoblastic hFOB 1.19 cellen en Osteosarcoom Saos-2 cellen. Osteosarcoom-afgeleide cellen worden vaak gebruikt als osteoblastic modellen en Saos-2 cellen zijn de meest volwassen osteoblastic teken19 bewaard terwijl ongedifferentieerde menselijke foetale hFOB cellen veel als een model voor de normale osteoblastic gebruikt worden differentiatie20. Hun mineralisatie profielen werden geanalyseerd door verschillende methoden: Alizarine Red-S (S-AR) kleuring, ultraviolet (UV) licht visualisatie, Transmissie Electronenmicroscopie (TEM) imaging, energie dispersieve x-stralen Microanalyse (EDX) kwantificatie en ion in kaart brengen. Het voordeel van de TEM-EDX over alternatieve technieken die worden gebruikt in eerdere studies is dat het kwantitatieve en kwalitatieve resultaten van ion vervanging in apatiet kristallen4,5,21geeft. Het algemene doel van het gebruik van de TEM-EDX was te vinden van een eenvoudige methode voor beeldbewerking en kwantificering van de verdeling van de Ca, F en Cl-ionen in verschillende mineralen uit verschillende soorten cellen tijdens de verschillende fasen van het proces van mineralisatie. Deze methode is met succes gebruikt, bijvoorbeeld voor het toezicht op de interactie van zink nanodeeltjes met naast elkaar bestaande chemische stoffen en hun gecombineerde effecten op aquatische organismen22. In een andere studie, een koperen photocatalyst op titanium materialen in waterige oplossing werd uitgebreid gekenmerkt door middel van inductief gekoppeld plasma optische-emissiespectrometrie (ICP-OES), N2 physisorption (inzet), XRD, UV-vis DRS, FT-IR, Raman spectroscopie, TEM-EDX en photoelectrochemical metingen23. Ons doel was om te vergelijken van de herkomst en eigenschappen van blaasjes en mineralen in twee cellijnen te begrijpen van het mechanisme waarmee mineralisatie tijdens werden differentiatie.
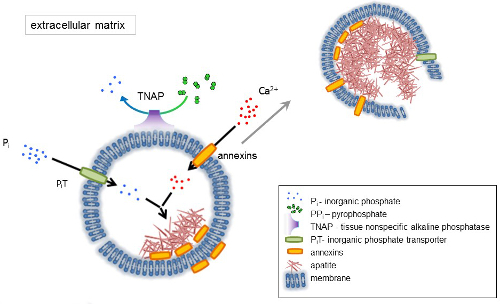
Figuur 1 . Regeling van de eerste stappen van mineralisatie in werden cellen waarbij de synthese van de extracellulaire matrix (ECM) eiwitten en vrijlating van matrix blaasjes (MVs) van de membraan. MVs accumuleren calcium door de werking van calcium bindende proteïnen, annexins en fosfaat, door de werking van een anorganisch fosfaat transporter (PiT) gevolgd door de activiteit van weefsel aspecifieke alkalisch fosfatase (TNAP), die dephosphorylates PPik om Pik, teneinde apatiet nucleatie. Vervolgens MVs desintegreren en vrij van apatites aan het extracellulaire medium. De mineralisatie is opgelopen door de constante aanvoer van Pik en Ca2 + in de extracellulaire middellange4,5. Klik hier voor een grotere versie van dit cijfer.
