Knochen ist eine Art von Bindegewebe besteht aus zwei Teilen: Bio (Zellen und Kollagenfasern) und Mineralien (Calcium und Phosphat-Verbindungen). Die mineralischen Hauptbestandteile in den Knochen sind Apatite1. Verschiedene Arten von Mineralisierung-kompetente Zellen in Knochen (Osteoblasten), Zähne (Odontoblasts) und im Knorpel (Chondrozyten) regulieren die ersten Schritte der Mineralisierung durch Produktion von Proteinen der extrazellulären Matrix (ECM) und loslassen matrix Vesikel (MVs) (Abbildung 1). MVs sind 100-300 nm Durchmesser Vesikel, die Calcium- und Phosphatspiegel, die Erleichterung der Apatit Keimbildung zu sammeln und anschließend binden an Kollagen2,3. Dann zerfallen MVs Apatite auf das extrazelluläre Medium lösen. Die Apatite weiterhin in Kontakt mit Kollagenfasern wachsen und bilden der Knochenmatrix. Die Mineralisierung wird durch die konstante Versorgung mit Pich und Ca2 + im extrazellulären Medium getragen. Einige kürzlich veröffentlichten Daten unterstützen unsere Modell-4,–5. Weichteile nicht unter physiologischen Bedingungen mineralisieren. Ektopische Verkalkung kann jedoch unter pathologischen Bedingungen wie Gefäßverkalkung3auftreten. Vaskuläre Zellen, die Osteoblasten Phänotyp erwerben können MVs produzieren, die Keimbildung von Apatite zu induzieren und Mineralisierung in den medialen und Intima Schichten der Wand der Blutgefäße zu initiieren. Seit ektopische Verkalkung ähneln normalen Endochondral Mineralisierung3, Verständnis der molekularen Mechanismen der Mineralisierung der Knochen Zellen und Chondrozyten sollen einige Hinweise auf ektopische Verkalkung der Weichteile, die gebildet.
Die Entwicklung des skelettartigen Gewebe wird durch verschiedene Enzyme, Wachstumsfaktoren und Promotoren oder Inhibitoren der Mineralisierung geregelt. Die Antagonistische Wirkung der Gewebe-unspezifische alkalische Phosphatase (TNAP) (Abbildung 1) und Ectonucleotide Pyrophosphatase/Phosphodiesterase kontrolliert ich (NPP1), zusammen mit Ankyrin (ANK), anorganisches Pyrophosphat (PPich) Konzentration 6. PPich, ein potenter Inhibitor der HA-Formation ist hydrolysiert durch TNAP; NPP1 hydrolysiert Nukleotid Triphosphate um PPich zu bilden, während ANK PPich in das ECM aus der Zelle exportiert. Das Verhältnis von Pi/PPi kann Apatit Bildung7,8 mit möglichen pathologischen folgen9regulieren.
Die MV-Membran ist in Ionen-Transport-Proteine angereichert, die die erste Ausfällung von Kalzium und Phosphat im Inneren der MVs bei der Keimbildung (Abbildung 1) zu erleichtern. Die Phosphat-Transporter 1 (PiT) hilft, um Pich erzeugt im Perivesicular Raum in der MVs10,11zu integrieren. Polymerasen können beteiligt sein, in die Bindung und den Transport von Ca2 + und in den biophysikalischen Prozess, der Mineralisierung in der MV Lumen12,13initiiert. Wir favorisieren die Hypothese vorgeschlagen, für die Mineralisierung innerhalb Intrazytoplasmatische Vesikel des internen Keimbildung von Apatit innerhalb der MV vor ihrer Ausbreitung in der ECM-14,–15. In-vitro- Modellierung bestätigt die Induktion von Ca2 +/ pich komplexe Bildung in Proteoliposomes aus PS und AnxA516hergestellt. Dies kann darauf hindeuten, dass Akkumulation von Ca2 +, Pich, AnxA5 und PS-komplexe in Lipid Rafts der Mikrovilli-wie Membranesrepresent der Keimbildung Kern (NC) Apatit innerhalb MVs. Polymerasen und TNAP besitzen auch Kollagen-Bindung Kapazitäten, die möglicherweise hilfreich bei der Platzierung von MVs entlang Kollagenfasern und bei der Förderung der Ausbreitung von Mineralisierung in der ECM. Fetuin-A und Osteopontin (OPN)17, sind bekannt als Inhibitoren der Apatit-Formation, die die Ausbreitung der Mineralisierung auf dem kollagenen Gerüst verlangsamen kann. Keimbildung und Vermehrung sind unterschiedliche Veranstaltungen, erstere vor dem letzteren, und sowohl für den Prozess der pathologischen Mineralisierung relevant sein können.
Um herauszufinden, wie die Chemie des Calcium-Phosphat-komplexen physiologischen Mineralisierung und ektopische Verkalkung ändern kann, ist es notwendig, die Mineralien produziert von Zellen zu identifizieren. Apatite sind eine Gruppe von Kalzium und Phosphat enthalten Mineralien mit dem allgemeinen Kristall Zelle Formel Ca10(PO4)6X2, wobei X = Cl, F, OH. Sie gelten als18folgt: Fluorapatit (FA) Ca10(PO4)6F2, Chlorapatite (CA) Ca10(PO4)6Cl2 und Hydroxylapatit (HA) Ca10(PO4 ),6(OH)2.
Die Wahl der Osteoblasten Zelllinien induzieren MINERALBILDUNG ist entscheidend, denn jede Zelllinie ein klares Profil der Mineralisierung zeigt. In diesem Bericht verglichen wir die Keimbildung der Mineralien von zwei ausgewählten menschlichen zellmodelle Mineralisierung: osteoblastischen hFOB 1.19 und Osteosarkom Saos-2 Zellen. Osteosarkom-abgeleitete Zellen werden häufig als osteoblastischen Modelle verwendet und Saos-2 Zellen haben die reifsten osteoblastischen Charakter19 beibehalten, während undifferenzierte menschlichen fetalen hFOB Zellen häufig als Modell für normale osteoblastischen verwendet werden Differenzierung-20. Ihre Mineralisierung Profile wurden mit verschiedenen Methoden analysiert: Alizarin rot-S (AR-S) Färbung, ultravioletten (UV) Licht Visualisierung Übertragung Elektronenmikroskopie (TEM) Bildgebung, Energy dispersive x-ray Mikroanalyse (EDX) Quantifizierung und Ion Mapping. Der Vorteil von TEM-EDX über alternative Techniken verwendet in früheren Studien ist, dass es quantitative und qualitative Ergebnisse der Ionen-Austausch in Apatit Kristalle4,5,21 gibt. Das übergeordnete Ziel der Verwendung von TEM-EDX war, eine einfache Methode für Bildgebung und Quantifizierung der Verteilung von Ca, F und Cl-Ionen in verschiedenen Mineralien aus verschiedenen Arten von Zellen während der Phasen des Prozesses Mineralisierung zu finden. Diese Methode erfolgreich, zum Beispiel dient zur Überwachung der Wechselwirkung von Zink-Nanopartikel mit verschiedenen Chemikalien und ihre kombinierte Wirkung auf Wasserorganismen22. In einer weiteren Studie, ein Kupfer Photocatalyst auf Titan-Materialien in wässriger Lösung ausführlich zeichnete sich durch optische emissionsspektrometrie induktiv gekoppelte Plasma (ICP-OES), N2 Physisorption (BET), XRD, UV-Vis-DRS, FT-IR, Raman Spektroskopie, TEM-EDX und photoelektrochemische Messungen23. Unser Ziel war es, die Herkunft und Eigenschaften der Vesikel und Mineralien in beiden Zelllinien, den Mechanismus zu verstehen, der Mineralisierung im knöchernen Differenzierung steuert zu vergleichen.
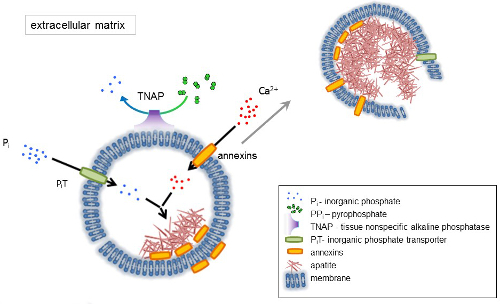
Abbildung 1 . Schema der ersten Schritte der Mineralisierung im knöchernen Zellen mit der Synthese von Proteinen der extrazellulären Matrix (ECM) und Freisetzung von Matrix Vesikel (MVs) aus der Membran. MVs akkumulieren Kalzium durch die Einwirkung von Kalzium-bindende Proteine, Polymerasen und Phosphat, durch das Wirken des ein anorganisches Phosphat-Transporter (PiT) gefolgt von der Aktivität der Gewebe-unspezifische alkalische Phosphatase (TNAP), dem dephosphorylates PPich Pich, wodurch Apatit Keimbildung. Dann MVs zerfallen und freigeben Apatite auf das extrazelluläre Medium. Die Mineralisierung wird durch die konstante Versorgung mit Pich und Ca2 + in der extrazellulären mittlere4,5getragen. Bitte klicken Sie hier für eine größere Version dieser Figur.
