Ben er en type bindevev består av to deler: organisk (celler og kollagen fibrene) og mineral (kalsium og fosfat forbindelser). Mineral hovedkomponentene i bein er direkte oversatt: apatitter1. Ulike typer mineralisering kompetente celler i bein (osteoblasts), tenner (odontoblasts) og brusk (chondrocytes) regulere forbokstaven skritt av mineralisering av produserer proteiner av den ekstracellulære matrisen (ECM) og slippe matrise blemmer (MVs) (figur 1). MVs er 100-300 nm diameter blemmer som akkumuleres kalsium og fosfat tilrettelegge apatitt nucleation og deretter binde kollagen2,3. Deretter nedbryte MVs for å løslate direkte oversatt: apatitter til ekstracellulære medium. De direkte oversatt: apatitter fortsette å vokse i kontakt med kollagen fibrene og danne bein matrix. Mineraliseringen er påført konstante tilførselen av Pjeg og Ca2 + i ekstracellulære medium. Noen nylig publiserte data støtte vår modell4,5. Myke vev ikke mineralize under fysiologiske forhold. Ektopisk forkalkninger kan imidlertid oppstå under patologiske tilstander som vaskulær forkalkninger3. Vaskulære celler som kjøper osteoblast fenotypen kan produsere MVs som induserer nucleation av direkte oversatt: apatitter og starte mineralisering i mediale og intimal lagene på veggen av blod fartøy. Siden ektopisk forkalkninger ligne normal endochondral mineralisering3, forstå molekylære mekanismer av mineralisering av osseous celler og chondrocytes bør gi noen hint om ektopisk forkalkning av mykt vev som er dannet.
Utvikling av skjelettet vev er regulert av ulike enzymer, vekstfaktorer, og arrangører og hemmere av mineralisering. Handlingen antagonistiske vev-uspesifikke alkalisk fosfatase (TNAP) (figur 1) og ectonucleotide pyrophosphatase/fosfodiesterase jeg (NPP1), sammen med ankyrin (ANK), styrer uorganiske pyrophosphate (PPjeg) konsentrasjon 6. PPjeg, en potent hemmer av HA formasjon, er hydrolyzed av TNAP; NPP1 hydrolyzes nucleotide triphosphates å danne PPjeg mens ANK eksporterer PPjeg fra cellen til ECM. Pi/PPT forholdet kan regulere apatitt formasjon7,8 med mulige patologisk konsekvenser9.
MV membranen er beriket i ion transport proteiner som letter første nedbør av kalsium og fosfat inne MVs under nucleation prosessen (figur 1). Den fosfat transporter 1 (PiT) bidrar til å innlemme Pjeg generert i perivesicular plass i MVs10,11. Annexins kan være involvert i bindingen og transport av Ca2 + og Biofysiske prosessen som starter mineralisering i MV lumen12,13. Vi ønsker hypotesen foreslått tidligere mineralisering innen Intracytoplasmatisk blemmer av interne nucleation av apatitt inne MV før sin forplantning i ECM14,15. In vitro modellering bekreftet induksjon av Ca2 +/Pjeg komplekser formasjon i proteoliposomes laget av PS og AnxA516. Dette kan tyde på at opphopning av Ca2 +, Pjeg, AnxA5 og PS komplekser i lipid flåter av microvilli-lignende membranesrepresent nucleation kjernen (NC) i apatitt Mvs Annexins og TNAP har kollagen-bindende kapasitet som kan være nyttig i å plassere MVs langs kollagenfibre og stimulere utbredelsen av mineralisering i ECM. Fetuin A og osteopontin (OPN)17, kalles hemmere av apatitt formasjon som kan forsinke spredning av mineralisering på collagenous skafottet. Begge kan være relevante for prosessen med patologisk mineralisering nucleation og overføring er forskjellige arrangementer, tidligere foran sistnevnte.
For å oppdage hvordan kjemien i kalsium fosfat komplekser kan endre fysiologiske mineralisering og ektopisk forkalkninger, er det nødvendig å identifisere mineraler produsert av cellene. Direkte oversatt: apatitter er en gruppe av kalsium og fosfat som inneholder mineraler med generelle krystall enheten cellen formelen Ca10(PO4)6X2, der X = Cl, F, OH. De er klassifisert som følger18: fluorapatite (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 og hydroxyapatite (HA) Ca10(PO4 )6(OH)2.
Valget av osteoblast linjer å skape mineral er viktig, siden hver celle linje viser en tydelig profil av mineralisering. I denne rapporten vi sammenlignet nucleation av mineraler av to valgte human celle modeller av mineralisering: osteoblastic hFOB 1,19 cellene og osteosarcoma Saos-2. Osteosarcoma-avledet celler brukes ofte som osteoblastic og Saos-2 celler har bevart de fleste eldre osteoblastic karakter19 mens udifferensierte menneskelige føtal hFOB celler er mye brukt som modell for vanlig osteoblastic differensiering20. Deres mineralisering profiler ble analysert av ulike metoder: Alizarin rød-S (AR-S) flekker, ultrafiolett (UV) lys visualisering, overføring elektronmikroskop (TEM) bildebehandling, energi dispersiv X-ray microanalysis (EDX) kvantifisering og ion tilordning. Fordelen med TEM-EDX over alternative teknikker som brukes i tidligere studier er at det gir kvantitative og kvalitative resultatene av ion erstatning i apatitt krystaller4,5,21. Det overordnede målet med å bruke TEM-EDX var å finne en enkel metode for bildebehandling og kvantifisering av fordelingen av Ca, F og Cl ioner i ulike mineraler fra forskjellige celler i forskjellige stadier av mineralisering. Denne metoden har blitt brukt, for eksempel for overvåking samspillet av sink nanopartikler coexisting kjemikalier og deres kombinerte effekten vannlevende organismer22. I en annen studie, en kobber photocatalyst på Titan materialer i vandig løsningen var mye preget med Induktivt kombinert plasma optisk utslipp massespektrometri (ICP-OES), N2 physisorption (innsats), XRD, UV-vis DRS, FT-IR, Raman spektroskopi og TEM-EDX photoelectrochemical målinger23. Vårt mål var å sammenligne opprinnelse og egenskaper av blemmer og mineraler i to linjer å forstå mekanismen som styrer mineralisering under osseous differensiering.
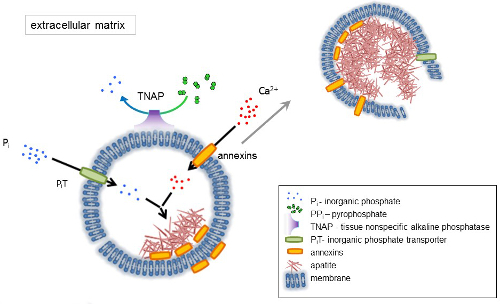
Figur 1 . Ordningen av de første trinnene av mineralisering i osseous celler syntesen av ekstracellulær matrix (EFM) proteiner og utgivelsen av matrix blemmer (MVs) fra membranen. MVs samle kalsium handlingen av kalsium bindende proteiner, annexins og fosfat, gjennom handling av en uorganisk fosfat transporter (PiT) etterfulgt av aktiviteten til vev ikke-spesifikk alkalisk fosfatase (TNAP), som dephosphorylates PPjeg Pjeg, og dermed tilrettelegge apatitt nucleation. Deretter MVs oppløsning og slipp direkte oversatt: apatitter til ekstracellulære medium. Mineraliseringen er påført konstante tilførselen av Pjeg og Ca2 + i ekstracellulære middels4,5. Klikk her for å se en større versjon av dette tallet.
