Кость — это тип соединительной ткани, состоящей из двух частей: органические (клетки и коллагеновые волокна) и минеральных (соединений кальция и фосфата). Основные минеральные компоненты в кости, Апатиты1. Различные типы минерализации компетентных клеток в кости (остеобласты), зубы (odontoblasts) и хряща (хондроцитов) регулируют первоначальные шаги минерализации, производя белков внеклеточного матрикса (ECM) и выпускать матрица везикулы (МВС) (рис. 1). МВС являются 100-300 Нм диаметр пузырьков, которые накапливаются кальция и фосфатов, содействия апатита нуклеации и впоследствии связать коллаген2,3. Затем MVs дезинтегрируют выпустить апатитов внеклеточных среды. Апатиты продолжают расти при контакте с коллагеновых волокон и формируют костной матрицы. Минерализация подкрепляется постоянные поставки P,я и Ca2 + в внеклеточных среды. Некоторые недавно опубликованные данные поддерживают нашу модель4,5. Мягких тканей не минерализации в физиологических условиях. Однако эктопической кальцификации может произойти в патологических условиях например сосудистой кальцификации3. Сосудистые клетки, которые приобретают остеобластов фенотипом могут производить MVs, которые вызывают нуклеации апатитов и инициировать минерализации в медиальной и интимы слои стенки кровеносных сосудов. Начиная с эктопической кальцификации напоминают нормальный endochondral минерализация3, понимание молекулярных механизмов минерализация костных клеток и хондроцитов следует обеспечить некоторые подсказки на Эктопическая кальцификация мягких тканей, которые являются сформирован.
Развитие скелетных тканей регулируется различные ферменты, факторы роста и промоутеров или ингибиторов минерализации. Антагонистическое действие ткани неспецифические щелочной фосфатазы (TNAP) (рис. 1) и ectonucleotide пирофосфатаза/фосфодиэстеразы я (NPP1), вместе с Анкирины (АНК), контролирует концентрации неорганических пирофосфат (PPя) 6. PPя, мощным ингибитором формирования HA, гидролизуется по TNAP; NPP1 гидролизует нуклеотидов трифосфаты сформировать PP,я пока ANK экспортирует PP,я из ячейки в ECM. Pi/PPi соотношение может регулировать апатита формирования7,8 возможных патологических последствий9.
Мембрана MV обогащается в ионный транспорт белков, которые облегчают начальный осадков кальция и фосфатов внутри векторы во время процесса нуклеации (рис. 1). Фосфат транспортер 1 (Пит) помогает включить Pя сгенерирована perivesicular пространства в МВС10,11. Аннексины могут быть вовлечены в привязке и транспорта Ca2 + и в процессе биофизических, который инициирует минерализации в MV просвета12,13. Мы выступаем за гипотеза, предложил ранее, для минерализации в интрацитоплазматической пузырьков внутреннего нуклеации апатита внутри мВ до его распространения в ECM14,15. В vitro моделирования подтвердил индукции формирования Ca2 +/ pя комплексов в proteoliposomes из16Л.С. и AnxA5. Это может означать, что накопление Ca2 +, P,я, AnxA5 и ПС комплексов в липидной плоты микроворсинки как membranesrepresent нуклеации ядро (NC) апатита в пределах Мпротив Аннексины и TNAP обладают также коллаген привязки возможности, которые могут быть полезны в размещении MVs вдоль волокон коллагена и в стимулировании распространения минерализации в ECM. Fetuin A и Остеопоэтин (OPN)17, известны как ингибиторы формирования апатита, которая может замедлить распространение минерализации на коллагеновых эшафот. Зарождения и распространения являются различные события, бывший до последнего, и оба могут быть актуальными для процесса патологических минерализации.
Чтобы узнать, как химический состав кальция фосфат комплексов могут изменить физиологических минерализации и эктопической кальцификации, это необходимо для идентификации минералов, производимые клеток. Апатиты группа кальция и фосфатов, содержащие минералы с общего кристалл блок клеток формулой Ca10(PO4)6X2, где X = Cl, F, OH. Они классифицируются следующим образом18: fluorapatite (ФА) Ca6F10(PO4)2, хлорапатитом (CA) Ca10(PO4)6Cl2 и гидроксиапатита (HA) Ca10(PO4 )6(OH)2.
Выбор остеобластов клеточных линий, чтобы побудить минеральные образования имеет решающее значение, поскольку в каждой ячейке строки экспонатов собственный профиль минерализации. В настоящем докладе, мы сравнили нуклеации минералов на две отдельных клеток человека модели минерализации: osteoblastic hFOB 1.19 клетки и клетки остеосаркома Saos-2. Остеосаркома, полученных клетки обычно используются как osteoblastic модели и Saos-2 клетки сохранили наиболее зрелой osteoblastic характер19 пока недифференцированные человеческого плода hFOB клетки широко используются как модель для нормальной osteoblastic дифференциация20. Их минерализации профили были проанализированы различными методами: пятнать ализарин красный-S (AR-S), ультрафиолетового (УФ) света визуализации, передачи электронной микроскопии (ТЕА) изображений, энергии энергодисперсионный рентгеновский микроанализ (EDX) количественный и Ион сопоставление. Преимущество ТЕА-EDX над альтернативные методы, используемые в предыдущих исследованиях является, что она дает количественные и качественные результаты замены иона в апатита кристаллов4,5,21. Общая цель использования ТЕА-EDX состояла в том, чтобы найти простой способ для создания образов и количественной оценки распределения ионов Ca, F и Cl в различных минералов из различных типов клеток на различных этапах процесса минерализации. Этот метод успешно используется, например, для контроля взаимодействия цинка наночастиц с сосуществующих химических веществ и их совокупное воздействие на водные организмы22. В другом исследовании, медные фотокатализатор на титановые материалы в водном растворе широко характеризуется с помощью индуктивно связанной плазмы оптической эмиссионной спектрометрии (ICP-OES), N2 physisorption (BET), Дифракционные, UV-vis DRS, FT-IR, Раман спектроскопия, ТЕА-EDX и фотоэлектрохимические измерения23. Нашей целью было сравнить происхождение и свойства везикулы и минералов в двух клеточных линий, чтобы понять механизм, который управляет минерализации при костной дифференциации.
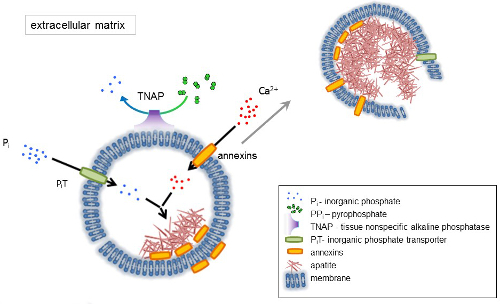
Рисунок 1 . Схема из первоначальных шагов минерализации в костных клеток, синтез белков внеклеточного матрикса (ECM) и выпуска Матричные везикулы (МВС) от мембраны. МВС накопления кальция через действия кальция связывания белков, Аннексины и фосфат, через действие неорганического фосфата транспортера (Пит) следуют активности тканей неспецифической щелочной фосфатазы (TNAP), который dephosphorylates PPя Pя, способствуя апатита зародышеобразования. Затем MVs распадаться и отпустите апатитов внеклеточных среды. Минерализация поддерживается постоянной поставке P,я и Ca2 + в внеклеточной средний4,5. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
