Ben är en typ av bindväv som består av två delar: ekologisk (celler och kollagenfibrer) och mineraler (kalcium och fosfat föreningar). Mineraliska huvudkomponenterna i ben är apatites1. Olika typer av mineralisering-kompetenta celler i skelettet (osteoblaster), tänder (odontoblaster) och brosk (kondrocyter) reglera de första stegen av mineralisering av producerar proteiner av den extracellulär matrixen (ECM) och släppa matris blåsor (MVs) (figur 1). MVs är 100-300 nm diameter blåsor som ackumuleras kalcium och fosfat underlätta apatit kärnbildning och därefter binda till kollagen2,3. Sedan, MVs upplösas för att släppa apatites till extracellulära medium. Apatites fortsätter att växa i kontakt med kollagenfibrer och bilda benmatrix. Mineraliseringen upprätthålls av den konstant tillförselen av Pjag och Ca2 + i det extracellulära mediet. Några nyligen publicerade data stödjer vår modell4,5. Mjuka vävnader inte mineralize under fysiologiska betingelser. Ektopisk förkalkning kan dock inträffa under sjukdomstillstånd såsom vaskulär förkalkning3. Vaskulära celler som förvärvar osteoblast fenotypen kan producera MVs som inducerar kärnbildning av apatites och initiera mineralisering i väggen mediala och intimans lager av blodkärl. Sedan ektopisk förkalkning likna normala endochondral mineralisering3, förstå de molekylära mekanismerna av mineralisering av bendefekter celler och kondrocyter bör ge några ledtrådar om ektopisk förkalkning av mjukvävnad som är bildas.
Utvecklingen av skelettet vävnader regleras av olika enzymer, tillväxtfaktorer, och initiativtagare eller hämmare av mineralisering. Åtgärden antagonistiska vävnad-ospecifik alkaliskt fosfatas (TNAP) (figur 1) och ectonucleotide pyrophosphatase/fosfodiesteras jag (NPP1), tillsammans med ankyrin (ANK), styr oorganisk pyrofosfat (PPjag) koncentration 6. PPjag, en potent hämmare av HA bildandet, hydrolyseras av TNAP; NPP1 hydrolyserar nukleotid trifosfater bildar PPjag medan ANK export PPjag från cellen till ECM. Förhållandet Pi/PPi kan reglera apatit bildandet7,8 med eventuella patologiska konsekvenserna9.
MV membranet är berikad i ion transportproteiner som underlättar inledande utfällning av kalcium och fosfat inuti MVs under processen kärnbildning (figur 1). Fosfat transportören 1 (PiT) hjälper till att införliva Pjag genereras i perivesicular utrymme till MVs10,11. Annexins kan vara inblandade i bindningen och transport av Ca2 + och biofysiska processen som initierar mineralisering i MV lumen12,13. Vi gynnar den hypotesen, föreslog tidigare, för mineralisering inom intracytoplasmatisk blåsor av interna kärnbildning av apatit inuti MV innan dess förökning i ECM14,15. In vitro modellering bekräftade induktion av Ca2 +/Pjag komplex bildande i proteoliposomes gjorda av PS och AnxA516. Detta kan tyda på att ansamling av Ca2 +, Pjag, AnxA5 och PS komplex i lipid rafts av Mikrovilli-liknande membranesrepresent kärnbildning kärnan (NC) i apatite inom MVs. Annexins och TNAP också äger kollagen bindande kapacitet som kan vara till hjälp i att placera MVs längs kollagenfibrer och stimulera spridningen av mineralisering i ECM. Fetuin A och osteopontin (OPN)17, är kända som hämmare av apatit bildandet som kan bromsa förökningen av mineralisering på kollagena schavotten. Kärnbildning och förökning är separata händelser, den förstnämnda som föregår den senare, och båda kan vara relevanta för processen av patologiska mineralisering.
För att upptäcka hur kemin av kalciumfosfat komplex kan ändra fysiologiska mineralisering och ektopisk förkalkning, är det nödvändigt att identifiera de mineraler som produceras av celler. Apatites är en grupp av kalcium och fosfat som innehåller mineraler med den allmänna crystal enhet cell formeln Ca10(PO4)6X2, där X = Cl, F, OH. De klassificeras enligt följande18: Fluorapatit (FA) Ca10(PO4)6F2, chlorapatite (CA) Ca10(PO4)6Cl2 och hydroxyapatit (HA) Ca10(PO4 )6(OH)2.
Valet av osteoblast cellinjer att framkalla mineral bildas är avgörande, eftersom varje cellinje uppvisar en tydlig profil av mineralisering. I detta betänkande, Vi jämförde kärnbildning av mineraler av två valda mänsklig cellmodeller av mineralisering: osteoblastiska hFOB 1.19 och osteosarkom Saos-2 celler. Osteosarkom-derived celler används ofta som osteoblastiska modeller och Saos-2-celler har bevarat de mest mogna osteoblastiska karaktär19 medan odifferentierade mänskliga fostrets hFOB celler används allmänt som en modell för normal osteoblastiska differentiering20. Sina mineralisering profiler analyserades av olika metoder: Alizarin Red-S (AR-S) färgning, ultraviolett (UV) ljus visualisering, överföring elektronmikroskopi (TEM) imaging, energy dispersive X-ray mikroanalys (EDX) kvantifiering och ion kartläggning. Fördelen med TEM-EDX jämfört med alternativa tekniker som används i tidigare studier är att det ger kvantitativa och kvalitativa resultat av ion ersättare i apatite kristaller4,5,21. Det övergripande målet med TEM-EDX var att hitta en enkel metod för avbildning och kvantifiering av fördelningen av Ca, F och Cl-joner i olika mineraler från olika typer av celler under olika faser av processen mineralisering. Denna metod har använts framgångsrikt, till exempel för att övervaka växelverkan av zink nanopartiklar med samtidiga kemikalier och deras kombinerade effekter på vattenlevande organismer22. I en annan studie, en koppar fotokatalysator på titanium material i vattenlösning präglades i stor utsträckning genom induktivt kopplad plasma OEC (ICP-OES), N2 physisorption (insats), XRD, UV-vis DRS, FT-IR, Raman spektroskopi och TEM-EDX photoelectrochemical mätningar23. Vårt syfte var att jämföra ursprung och egenskaper av blåsor och mineraler i två cellinjer att förstå mekanismen som styr mineralisering under bendefekter differentiering.
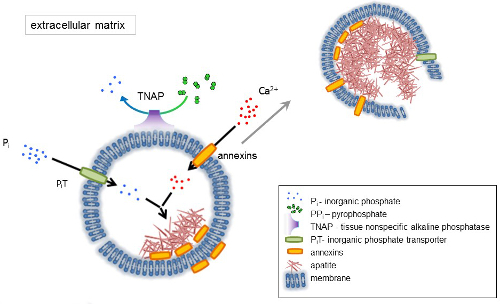
Figur 1 . Systematiken i de första stegen av mineralisering i bendefekter celler med syntesen av extracellulär matrix (ECM) proteiner och frisättning av matrix blåsor (MVs) från membranet. MVs ackumuleras kalcium genom åtgärder av kalcium bindande proteiner, annexins och fosfat, genom åtgärder av ett oorganiskt fosfat transportör (PiT) följt av aktiviteten av vävnad ospecifika alkaliska fosfataser (TNAP), som defosforylerar PPjag att Pjag, därigenom underlätta apatit kärnbildning. Sedan MVs upplösas och släppa apatites till extracellulära medium. Mineraliseringen upprätthålls av den konstant tillförselen av Pjag och Ca2 + i den extracellulära medium4,5. Klicka här för att se en större version av denna siffra.
