カスパーゼは、システインアスパラギン酸プロテアーゼのファミリーであり、イニシエーターカスパーゼおよび死刑執行人カスパーゼにグループ化することができる。死刑執行人カスパーゼは、カスパーゼ-3、-6および-7を含む。それらは二量体として細胞内に自然に見出され、アポトーシス1を実行するために開始カスパーゼによって切断される。開始カスパーゼには、ヒトカスパーゼ−1、−2、−4、−5、−8、−9、−10および−12が含まれる。それらは、近接誘導二量体化によって活性化され、自己タンパク質分解切断によって安定化される不活性ザイモゲン(プロカスパーゼ)として見出される2,3。炎症性カスパーゼは、開始カスパーゼ2のサブセットであり、ヒトにおけるカスパーゼ−1、−4、−5、および−12、ならびにマウス4、5におけるカスパーゼ−1、−11、および−12を包含する。アポトーシスの役割ではなく、炎症において中心的な役割を果たします。それらは、病原性侵入者8,9に応答して放出される最初のサイトカインであるプロインターロイキン(IL)−1βおよびプロIL−18 6,7のタンパク質分解的プロセシングおよび分泌を媒介する。カスパーゼ-1は、その活性化プラットフォームへの募集時に活性化される。インフラマソームと呼ばれる大きな分子量のタンパク質複合体(図1A)10。カスパーゼ−4、−5、および−11の二量体化は、非正準インフラマソーム経路11、12を介してこれらのプラットフォームとは独立して起こる。
正準インフラマソームは、インフラマソームセンサータンパク質、アダプタータンパク質ASC(CARDを含むアポトーシス関連スペック様タンパク質)、およびエフェクタータンパク質カスパーゼ-110からなる細胞質ゾル多量体タンパク質複合体である。最もよく研究された正準インフラマソームは、ピリンドメイン(NLRP)、NLRP1およびNLRP3を含むNOD様受容体ファミリー、CARD(NLRC)、NLRC4を含むNLRファミリー、および黒色腫2に存在しない(AIM2)である。それらはそれぞれ、ピリンドメイン、CARD、またはその両方のドメインを含む。CARD ドメインは、CARD を含むカスパーゼとそのアップストリーム アクティベーター間の相互作用を仲介します。したがって、N末端ピリンドメイン(PYD)とC末端CARDモチーフ13,14とからなる足場分子ASCは、NLRP110、NLRP3 15、およびAIM216インフラマソームへのカスパーゼ-1のリクルートに必要である。
各インフラマソームは、明確な炎症促進性刺激を認識する独自のセンサータンパク質にちなんで名付けられました(図1B)。この経路の活性化因子は、正準刺激と呼ばれる。インフラマソームは、微生物成分および組織ストレスのセンサとして機能し、炎症性カスパーゼ17の活性化を介して堅牢な炎症反応を誘発するように組み立てられる。インフラマソームアセンブリは、カスパーゼ−1活性化を開始し、その主要基質であるプロ−IL−1βおよびプロIL−18の成熟および分泌を媒介する。このプロセスは、2段階のメカニズムを介して行われます。第1に、プライミング刺激は、NF−κB経路の活性化を介して、ある種のインフラマソームタンパク質およびプロIL−1βの発現をアップレギュレートする。第二に、細胞内(正準)刺激は、プロカスパーゼ-1 6,7のインフラマソーム集合および動員を誘導する。
カスパーゼ−4およびカスパーゼ−5は、マウスカスパーゼ−11 11のヒトオルソログである。それらは、グラム陰性菌18、19、20の外膜に見られる分子である細胞内リポ多糖(LPS)および赤血球溶血21の産物である細胞外ヘムによって、インフラマソーム非依存的に活性化される。LPSがこれらのタンパク質のCARDモチーフに直接結合し、それらのオリゴマー化を誘導することが提案されている20。カスパーゼ-4またはカスパーゼ-5の活性化は、孔形成タンパク質ガスデルミンD(GSDMD)の切断を介してピロトーシスと呼ばれる炎症型の細胞死を誘導することによってIL-1β放出を促進する18,19。さらに、カスパーゼ−4およびGSDMD媒介性パイロプトーシス死から生じるカリウムイオンの流出は、NLRP3インフラマソームの活性化およびその後のカスパーゼ−1の活性化を誘導する22,23。したがって、カスパーゼ−4、−5、および−11は、特定の刺激に応答してパイロプトーシスおよびカスパーゼ−1活性化を誘導することができるLPSの細胞内センサと考えられる11、24。
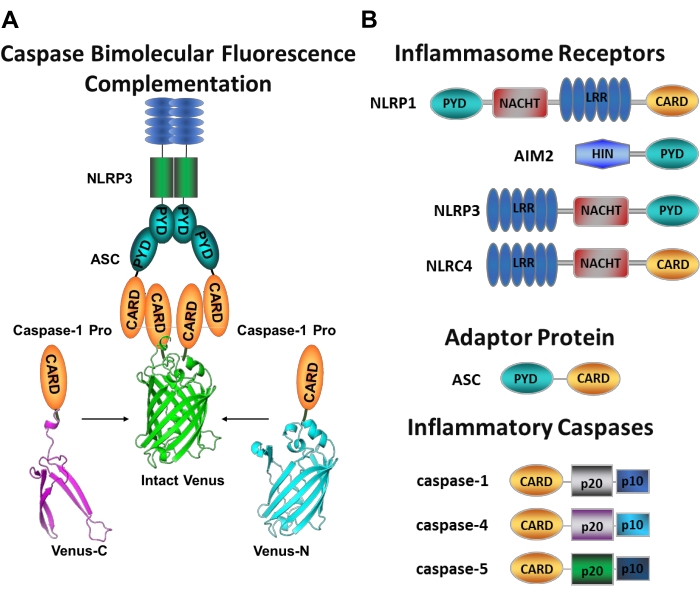
図1:炎症性カスパーゼおよびカスパーゼ-二分子蛍光相補(BiFC)アッセイ。(A)カスパーゼ-BiFC系を示す図では、金星の各非蛍光断片(Venus-CまたはVenus-N)に連結された2つのカスパーゼ-1プロドメイン(C1-pro)がNLRP3活性化プラットフォームにリクルートされ、金星にリフォールドと蛍光が強制されます。この複合体は、顕微鏡下で緑色の斑点として現れ、開始カスパーゼ活性化の最初のステップである炎症性カスパーゼ誘発近接の読み出しとして機能する。(b)インフラマソーム成分および炎症性カスパーゼのドメイン組織を示す模式図。この図の拡大版を表示するには、ここをクリックしてください。
特定の開始剤カスパーゼ活性化を測定することは困難であり、イメージングアプローチによってこれを行うために利用可能な方法は多くない。カスパーゼ二分子蛍光相補(BiFC)は、生細胞における炎症性カスパーゼ活性化を直接可視化するために使用できます(図1A)25。この技術は、最近、ヒト単球由来マクロファージ(MDM)21における使用に適応されている。カスパーゼBiFCは、炎症性カスパーゼ活性化における第1ステップを測定し、二量体化を促進するために近接を誘導する。CARD含有カスパーゼプロドメインをコードするプラスミドの発現は、光線上黄色蛍光タンパク質Venusの非蛍光断片に融合(Venus−C[VC])およびVenus−N[VN])が使用される。2つのカスパーゼプロドメインが活性化プラットフォームにリクルートされるか、または誘導された近接を受けると、金星の2つの半分が近接し、リフォールドおよび蛍光を発することを余儀なくされる( 図1A、B参照)。これは、特定の炎症性カスパーゼ活性化のリアルタイム読み出しを提供する。
ヒトMDMは、危険シグナルや病原体産物を同定するインフラマソーム遺伝子やパターン認識受容体を豊富に発現しています。これは、炎症性カスパーゼ経路の尋問に理想的な細胞型を提供する。さらに、それらは末梢血から、さらには患者サンプルから誘導して、特定の疾患状態における炎症性カスパーゼ活性化を評価することができる。このプロトコルでは、ヌクレオフェクションを使用してBiFCカスパーゼレポーターをMDMに導入する方法、エレクトロポレーションベースのトランスフェクション法、炎症性カスパーゼ活性化を誘導するために細胞を治療する方法、および顕微鏡アプローチを使用して活性カスパーゼ複合体を視覚化する方法について説明しています。さらに、この方法論は、これらの複合体の分子組成、細胞内局在化、動態、およびこれらの高度に秩序付けられた構造のサイズを決定するために適合させることができる25、26、27。
