Las caspasas son una familia de proteasas de cisteína aspartato que se pueden agrupar en caspasas iniciadoras y caspasas verdugo. Las caspasas verdugosas comprenden caspasa-3, -6 y -7. Se encuentran naturalmente en las células como dímeros y son escindidos por las caspasas iniciadoras para ejecutar la apoptosis1. Las caspasas iniciadoras incluyen caspasa humana-1, -2, -4, -5, -8, -9, -10 y -12. Se encuentran como zimógenos inactivos (pro-caspasas) que se activan por dimerización inducida por proximidad y se estabilizan mediante escisión autoprotolítica 2,3. Las caspasas inflamatorias son un subconjunto de las caspasas iniciadoras2 y abarcan caspasas-1, -4, -5 y -12 en humanos, y caspasa-1, -11 y -12 enratones 4,5. En lugar de un papel apoptótico, juegan un papel central en la inflamación. Median el procesamiento proteolítico y la secreción de pro-interleucina (IL)-1β y pro-IL-18 6,7, que son las primeras citoquinas que se liberan en respuesta a invasores patógenos 8,9. Caspasa-1 se activa al reclutar en su plataforma de activación; un complejo proteico de gran peso molecular denominado inflamasoma (Figura 1A)10. La dimerización de la caspasa-4, -5 y -11 ocurre independientemente de estas plataformas a través de una vía inflamasómica no canónica 11,12.
Los inflamasomas canónicos son complejos de proteínas multiméricas citosólicas que consisten en una proteína sensora de inflamasoma, la proteína adaptadora ASC (proteína similar a una mota asociada a la apoptosis que contiene una CARD) y la proteína efectora caspasa-110. Los inflamasomas canónicos más estudiados son la familia de receptores similares a NOD que contienen un dominio de pirina (NLRP), NLRP1 y NLRP3, la familia NLR que contiene una CARD (NLRC), NLRC4 y la ausente en el melanoma 2 (AIM2). Cada uno de ellos contiene un dominio de pirina, un CARD o ambos dominios. El dominio CARD media la interacción entre las caspasas que contienen CARD y sus activadores ascendentes. Por lo tanto, la molécula de andamio ASC, que se compone de un dominio de pirina N-terminal (PYD) y un motivo CARD C-terminal 13,14, es necesaria para el reclutamiento de caspasa-1 a los inflamasomas NLRP1 10, NLRP315 y AIM216.
Cada inflamasoma lleva el nombre de su proteína sensora única que reconoce distintos estímulos proinflamatorios (Figura 1B). Los activadores de esta vía se denominan estímulos canónicos. Los inflamasomas sirven como sensores para los componentes microbianos y el estrés tisular, y se ensamblan para desencadenar una respuesta inflamatoria robusta a través de la activación de las caspasas inflamatorias17. El ensamblaje del inflamasoma inicia la activación de la caspasa-1 para mediar la maduración y secreción de sus principales sustratos pro-IL-1β y pro-IL-18. Este proceso se produce a través de un mecanismo de dos pasos. En primer lugar, un estímulo de cebado regula al alza la expresión de ciertas proteínas inflamasomas y pro-IL-1β a través de la activación de la vía NF-κB. En segundo lugar, un estímulo intracelular (canónico) induce el ensamblaje del inflamasoma y el reclutamiento de procaspasa-1 6,7.
Caspasa-4 y caspasa-5 son los ortólogos humanos de la caspasa murina-1111. Se activan de manera independiente del inflamasoma por el lipopolisacárido intracelular (LPS), una molécula que se encuentra en la membrana externa de la bacteria Gram-negativa18,19,20, y por el hemo extracelular, un producto de la hemólisis de glóbulos rojos21. Se ha propuesto que el LPS se une directamente al motivo CARD de estas proteínas e induce su oligomerización20. La activación de la caspasa-4 o caspasa-5 promueve la liberación de IL-1β al inducir una forma inflamatoria de muerte celular llamada piroptosis a través de la escisión de la proteína formadora de poros gasdermin D (GSDMD)18,19. Además, el eflujo de iones potásicos resultantes de la caspasa-4 y la muerte piroptótica mediada por GSDMD induce la activación del inflamasoma NLRP3 y la posterior activación de la caspasa-1 22,23. Por lo tanto, la caspasa-4, -5 y -11 se consideran sensores intracelulares para LPS que son capaces de inducir la activación de la piroptosis y la caspasa-1 en respuesta a estímulos específicos11,24.
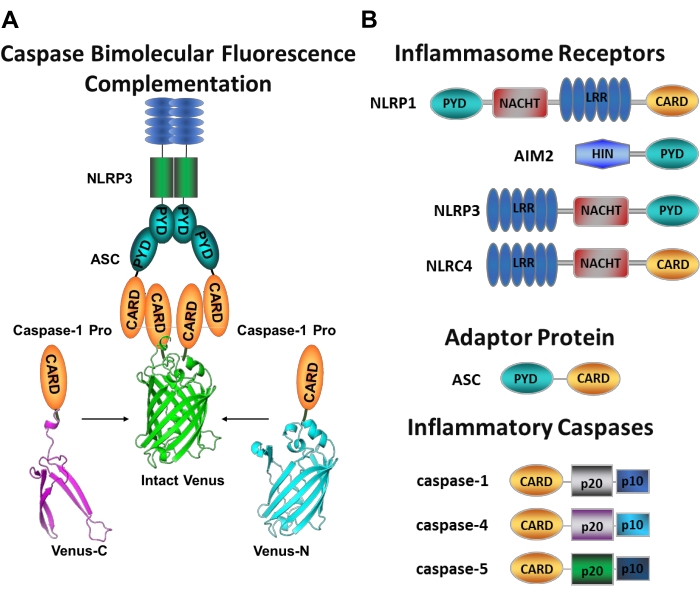
Figura 1: Ensayo de caspasas inflamatorias y complementación de fluorescencia caspasa-bimolecular (BiFC). (A) Diagrama que muestra el sistema caspasa-BiFC, donde dos prodominios de caspasa-1 (C1-pro) unidos a cada fragmento no fluorescente de Venus (Venus-C o Venus-N) se reclutan para la plataforma de activación NLRP3, obligando a Venus a replegarse y fluorescenciar. Este complejo aparece como una mancha verde bajo el microscopio y sirve como una lectura para la proximidad inflamatoria inducida por la caspasa, que es el primer paso en la activación de la caspasa iniciadora. (B) Esquema que muestra la organización del dominio de los componentes del inflamasoma y las caspasas inflamatorias. Haga clic aquí para ver una versión más grande de esta figura.
Medir la activación específica de las caspasas iniciadoras es difícil, y no hay muchos métodos disponibles para hacerlo mediante enfoques de imágenes. La complementación de fluorescencia bimolecular de caspasa (BiFC) se puede utilizar para visualizar la activación de la caspasa inflamatoria directamente en células vivas (Figura 1A)25. Esta técnica ha sido recientemente adaptada para su uso en macrófagos derivados de monocitos humanos (MDM)21. Caspasa BiFC mide el primer paso en la activación de la caspasa inflamatoria, proximidad inducida para facilitar la dimerización. Se utiliza la expresión de plásmidos que codifican el prodominio caspasa que contiene CARD fusionado con fragmentos no fluorescentes de la proteína fluorescente amarilla fotostable Venus (Venus-C [VC]) y Venus-N [VN]). Cuando los dos prodominios de caspasa se reclutan en su plataforma de activación o experimentan proximidad inducida, las dos mitades de Venus se acercan y se ven obligadas a replegarse y fluorescenciar (ver Figura 1A, B). Esto proporciona una lectura en tiempo real de la activación específica de la caspasa inflamatoria.
Los MDM humanos expresan abundantemente genes inflamasomas y receptores de reconocimiento de patrones que identifican señales de peligro y productos patógenos. Esto proporciona un tipo de célula ideal para el interrogatorio de las vías inflamatorias de la caspasa. Además, pueden derivarse de sangre periférica e incluso de muestras de pacientes para evaluar la activación de la caspasa inflamatoria en un estado de enfermedad específico. Este protocolo describe cómo introducir a los reporteros de caspasa BiFC en MDM utilizando nucleofection, un método de transfección basado en electroporación, cómo tratar las células para inducir la activación inflamatoria de la caspasa y cómo visualizar los complejos activos de caspasa utilizando enfoques de microscopía. Además, esta metodología se puede adaptar para determinar la composición molecular de estos complejos, la localización subcelular, la cinética y el tamaño de estas estructuras altamente ordenadas 25,26,27.
