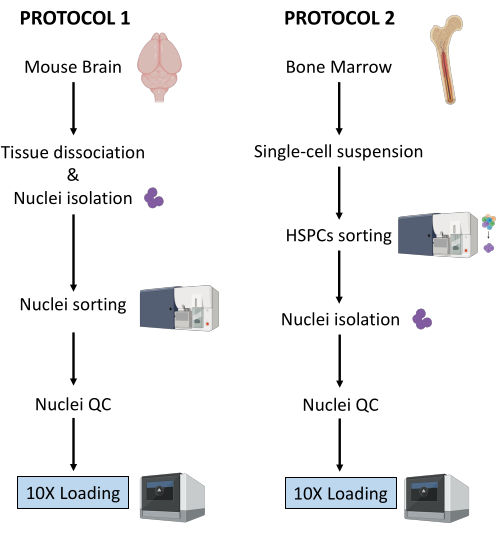
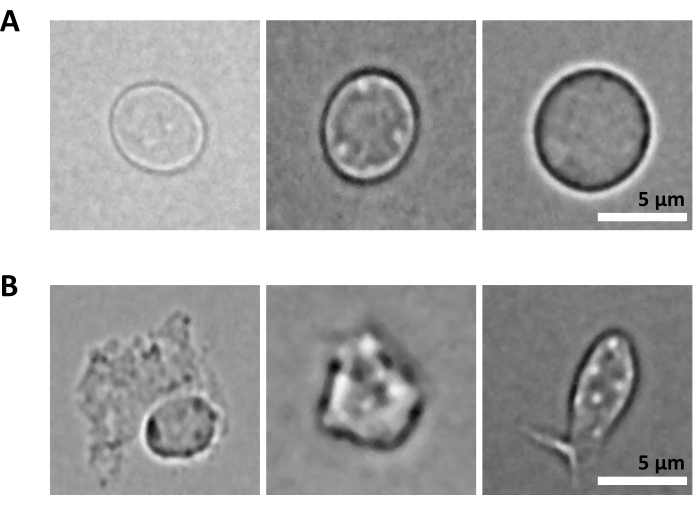
הליכי הניסוי נערכו תחת ציות רגולטורי קפדני לפרוטוקולים שאושרו על ידי ועדת האתיקה בניסויים בבעלי חיים (CETEA). לבידוד גרעיני המוח נעשה שימוש בעכברי C57BL/6 בני 3 חודשים. לצורך בידוד מח העצם נעשה שימוש בנקבות עכברי C57BL/6J בנות 8 שבועות במשקל 18 גרם. 1. טיהור גרעינים ממוח העכבר הערה: יש ללבוש כפפות לטקס או ניטריל בכל עת במהלך ההליך. מומלץ מאוד ששני אנשים יבצעו את הניסוי, ששלבים 1 עד 3 (כלומר, הכנת תרחיף הגרעינים היחיד) יבוצעו על ידי אדם אחד, ושלב 4 (כלומר, הכנת המיון) יבוצעו במקביל על ידי אדם אחר. מכיוון שהפרוטוקול רגיש מאוד לזמן, קריטי למזער את זמן עיבוד הדגימה על ידי הכנת הממיין מיד עם הכנת מתלה הגרעינים היחיד. הכנת ריאגנטים וחומריםבזהירות לעקר את כלי הדיסקציה על ידי autoclave (ב 121 ° C במשך 20 דקות) ולשטוף אותם עם אתנול 70% ממש לפני השימוש. הכינו צלחת פטרי אחת בכל דגימה, מלאה ב-2-3 מ”ל של מלח חוצץ פוספט (DPBS) קר כקרח (DPBS). מצננים את המיקרוצנטריפוגה ל -4 מעלות צלזיוס, ממלאים דלי בקרח, ומניחים את הזכוכית מקפיצה הומוגנייזר על קרח. הכן חיץ ליזה גרעיני על ידי הוספת digitonin לריכוז סופי של 0.01%, 10 מ”ל לדגימה. הכן את מאגר הצביעה על ידי הוספת מעכבי RNase למאגר צביעת התא לקבלת ריכוז סופי של 0.2 U/μL, 20 מ”ל לדגימה. הכן DPBS 0.04% BSA על ידי הוספת מעכבי RNase לריכוז סופי של 0.2 U/μL, 2 מ”ל לדגימה. הכינו 1 מ”ל של חיץ גרעינים מדולל בהתאם לפרוטוקול Multiome25. שמור את כל הריאגנטים והדגימות על קרח. דיסקציה של רקמותלהקריב עכברים באמצעות פרוטוקולים שאושרו על ידי המוסד. בפרוטוקול זה, ראשם של העכברים נערף לאחר מנת יתר של קטמין/קסילזין. חתכו את ראש העכבר במספריים והוציאו את המוח מהגולגולת כמתואר ב-Meyerhoff et al.26. העבירו מיד את המוח לצלוחית פטרי שהוכנה עם DPBS 1x קר כקרח תחת מיקרוסקופ סטריאופוני פולט אור (LED). חתכו את רקמת המוח בעזרת אזמל כדי להפריד בין אזורי עניין במוח (למשל, קליפת המוח האנטורינלית, ההיפוקמפוס, קליפת המוח הקדם-מצחית) והעבירו כל אזור לצלחת פטרי נפרדת המכילה DPBS 1x קר כקרח. שמור על קרח. בעזרת אזמל, לטחון את הרקמה לחתיכות <0.5 ס"מ כדי להקל על הומוגניזציה בשלב הבא. בעזרת מיקרופיפטה P1000, העבירו את הרקמה הטחונה ואת ה-DPBS 1x מצלחת הפטרי לצינור של 1.5 מ”ל. הקפידו להשתמש בצינורות העשויים מפלסטיק בעל קישור נמוך לחלבון. אפשרו לחתיכות הרקמה להיפרד על ידי כוח הכבידה. בזהירות להסיר את עודף של 1x DPBS באמצעות micropipette P1000.הערה: לאחר שלב זה, ניתן להקפיא את הרקמה הטחונה על ידי העברת צינורות החלבון בעלי הקישור הנמוך לקרח יבש ולאחר מכן לאחסן ב -80 ° C עד להמשך בידוד גרעינים. בידוד גרעיניםמלאו את מדף הזכוכית ב-2 מ”ל של חיץ גרעינים קרים כקרח בליזיס עם 0.01% דיגיטונין. מוסיפים את חתיכות הטישו בקפיצה.הערה: אם אתה עובד עם רקמה קפואה טרייה, הוסף את הרקמה הקפואה הטחונה ישירות למאגר הגרעינים ליזה 0.01% דיגיטונין; אל תתנו לרקמה להפשיר לפני. הומוגניזציה באמצעות זכוכית להקפיץ רקמות homogenizer 25 פעמים עם pestle A ולאחר מכן 25 פעמים עם pestle B. להעביר את homogenate לתוך צינור 15 מ”ל. הוסיפו עוד 2 מ”ל של חיץ גרעינים קרים כקרח עם 0.01% דיגיטונין ודגרו על קרח למשך 5 דקות. גרעיני צנטריפוגות ב 500 x גרם במשך 5 דקות ב 4 ° C. הסר את supernatant עם micropipette ולהוסיף 4 מ”ל של חיץ גרעינים קר כקרח ליזה עם 0.01% digitonin. דוגרים על קרח במשך 5 דקות ומסננים דרך מסננת תאים בגודל 40 מיקרומטר. גרעיני צנטריפוגות ב 500 x גרם במשך 5 דקות ב 4 ° C ולהסיר את supernatant עם micropipette. הוסף 4 מ”ל של חיץ צביעה כדי לשטוף את הגרעינים והצנטריפוגה ב 500 x גרם במשך 5 דקות ב 4 ° C. הסר את supernatant עם micropipette ו resuspend את הגלולה ב 4 מ”ל של חיץ צביעה. סנן דרך מסננת תאים 40 מיקרומטר וצנטריפוגה ב 500 x גרם במשך 5 דקות ב 4 ° C. השעיה מחדש ב 1 מ”ל של PBS עם 0.04% BSA. ספירת גרעינים כדי להבטיח עקביות של הכנת רקמות/גרעינים בין דגימות שונות. הוא צפוי לקבל ספירות גרעינים דומות מאותם אזורי מוח:הוסף 10 μL של 0.4% טריפאן כחול לצינור ריק של 0.5 מ”ל. מוסיפים 10 μL של הגרעינים ומערבבים 5x על ידי pipetting. ספור גרעינים באמצעות מונה תאים אוטומטי בהתאם להמלצות הספק. שמור את הגרעינים על קרח. מכינים גרעינים למיון.הערה: הגרעינים שחולצו משלבים 7-AAD, וצביעה זו משמשת לטיהורם על ידי ממיין תאים המופעל על ידי פלואורסצנטיות (FACS).העבר 100 μL של גרעינים לצינור FACS לבקרה לא מוכתמת. הוסף 10 μL של 7-AAD לגרעינים הנותרים, ולשמור 5 דקות על 4 ° C. מיין לפחות 0.5 x 106 גרעינים לפי FACS כדי לסלק כפילויות ולכלוך. מיון גרעינים באמצעות FACSהערה: בעוד שניתן לבצע מיון גרעינים במגוון רחב של ממייני תאים, הליך השימוש במכשירי BD FACSAria Fusion או BD FACSAria III מתואר כאן. מומלץ מאוד שהכיול וההתקנה של ממיין התאים יבוצעו תחת פיקוח, או על ידי משתמש מנוסה במכשיר. כדי לקצר את זמן עיבוד הדגימה, קריטי להכין את הממיין ברגע שמתלה הגרעינים הבודדים מוכן.כיול מכשיר FACSהפעל את סדרן התאים ואת המחשב. לאחר חיבור התוכנה למכשיר, הפעל את הליך ההפעלה הנוזלי. בחר Cytometer > Fluidic startup-up בתפריט הראשי ובצע את ארבעת השלבים. לחץ על בוצע לאחר השלמת כל אחד מהם. הכנס את פיית 70 מיקרומטר, הפעל את הזרם, והשאר את הזרם להתייצב למשך 15 דקות. התאם את המשרעת כדי לקבל היווצרות טיפה ולחץ על נקודה מתוקה. שים את מסנן הצפיפות הניטרלית (N.D) 1.0 ופתח את ממשק ההגדרה והמעקב אחר ציטומטר (CST). בקרת איכות יומית: לדלל חרוזי CST בתווך FACS (ראה המלצות ספק) ולבצע בקרת CST. לאחר השלמתו, החלף את N.D 1.0 ב- N.D 2.0. לדלל Accudrops בתווך FACS (ראה המלצות ספק) ולבצע עיכוב נפילה כמתואר בשלבים 6 עד 10. בתבנית הניסוי, בחר את הניסוי Accudrop Drop Delay ופתח את פריסת המיון עבור הצינור. בתוך חלון המצלמה התחתון, לחץ על מתח ולאחר מכן על מסנן אופטי כדי לאפשר הפעלת מטען על לוחות ההסטה ושימוש במסנן אופטי ספציפי מול המצלמה. ודא כי הרביע בצד ימין מציין 100. במידת הצורך, כוונן את בורג הלייזר האדום כדי למטב את פגיעת הלייזר. התאם את קצב הזרימה כדי להגיע למהירות של 1,000 עד 3,000 אירועים בשנייה. לחץ על מיין ובטל. ודא שהרביע השמאלי שווה ל- 100, והרביע הימני הוא 0. אם הרביע השמאלי נמוך מ- 95, בצע עיכוב אוטומטי. לחץ על מתח ולאחר מכן בדוק מיון. בקרה על איכות הפקדת הזרמים הצדדיים בצינורות האיסוף. במידת הצורך, התאימו את מיקום הזרמים הצדדיים באמצעות הזזת המחוונים. הגדרת מכשיר FACS למיון גרעינים.התחל את רכישת הגרעינים הבלתי מוכתמים. אלה משמשים להגדרת פיזור קדימה וצד, ואת מתח הגלאי עבור הפרמטר 7-ADD. הגדר את הפרמטרים כך שהאות 7-AAD של המדגם הלא מוכתם ייפול בתוך העשור הראשון של סולם היומן בתרשים הנקודות. התחל לרכוש את הצינור של גרעינים מוכתמים ב-7-AAD והגדר את אוכלוסיות הגרעינים באמצעות אסטרטגיית gating המבוססת על (1) FSC-A/SCC-A ולאחר מכן FSC-H/SSC-H עבור גודל וגרעיניות, (2) FSC-H/FSC-A עבור אפליית כפילויות, ו-(3) SSC-A/7-AAD עבור גרעינים חיוביים של 7-AAD (ראה איור 2A). ודא שהזרם והסטייה יציבים. במצלמת הזרם הצדדי, הפעל את מיון הבדיקה, הפעלת מתח ואשר את מיון הטיפות המדויק בצינור 1.5 מ”ל המותקן בצד שמאל. בחלון פריסת מיון , בחר את אוכלוסיית העניין, כפי שהוגדרה בשלב 2 (לעיל). באירועי יעד, בחר את הסף ברציף כדי לקבל לפחות 0.5 x 106 גרעינים לדגימה. תחת Precision, בחר 4-Way Purity. לאחר שתהיה מוכן, לחץ על מיין ואישור כדי להתחיל במיון גרעינים. בקרת איכות וספירה של גרעינים מטוהריםהערה: שלב זה ייעשה רק במהלך ניסוי הפיילוט לאופטימיזציה של שלבי הכנת הדגימה, במטרה לבדוק את טוהר הגרעינים הממוינים שיוטענו על שבב הכרום 10X. לאחר אופטימיזציה מלאה של הפרוטוקול, לא מומלץ לבצע שלב בקרת איכות זה בניסויי המעקב כדי למנוע בזבוז מיותר של גרעינים שנאספו שעשויים להיות זמינים במספרים נמוכים.בקרת טוהר על ידי ציטומטריית זרימההעבר 10 μL של הגרעינים הממוינים לתוך צינור FACS חדש המכיל 90 μL של DPBS עם 2% סרום בקר עוברי מומת בחום (HI-FBS). רכוש ותעד נתונים לאחר המיון כדי לאמת את טוהר המיון ואת הכדאיות. ודא שלפחות 98% מהגרעינים מופיעים בשער הריבית, כהגדרתו ב-4.2 (ראה איור 2B). ספירת הגרעינים המטוהריםצנטריפוגות ממוינות גרעינים במשך 5 דקות ב 500 x גרם וב 4 ° C בזהירות להסיר לחלוטין את supernatant באמצעות micropipette. השהה מחדש ב 100 μL של מאגר גרעינים מדולל. הוסף 10 μL של 0.4% טריפאן כחול לצינור ריק של 0.5 מ”ל. מוסיפים 10 μL מהגרעינים הממוינים ומערבבים פי 5 על ידי פיפטציה. ספור גרעינים באמצעות מונה תאים אוטומטי בהתאם להמלצות הספק. כוונן את ריכוז הגרעינים ל 3.5 x 106/mL, כלומר, 16,000 גרעינים לכל 5 μL. בקרת איכות של גרעינים מטוהרים באמצעות מיקרוסקופהערה: שלב זה ייעשה רק במהלך ניסוי הפיילוט לאופטימיזציה של שלבי הכנת הדגימה לבדיקת איכות הגרעינים שיוטענו על שבב הכרום 10X. לאחר אופטימיזציה מלאה של הפרוטוקול, לא מומלץ לבצע שלב בקרת איכות זה בניסויי המעקב כדי למנוע בזבוז מיותר של גרעינים שנאספו שעשויים להיות זמינים במספרים נמוכים.ודא שהחלקות המיקרוסקופ והחלקות הכיסוי נקיות ונטולות אבק. במידת הצורך, שטפו ושטפו את הכיסויים באתנול מוחלט וייבשו אותם במגבונים ללא מוך. מפזרים 25 μL של פולי-ל-ליזין בבארות המגלשה שישמשו וידגרור במשך 10 דקות בטמפרטורת החדר (RT), מוגן מפני אבק. הסר את עודף פולי-L-ליזין ולהוסיף 10 μL של השעיית גרעינים מטוהרים. יש לדגור במשך 5 דקות ב-RT, מוגן מאבק. הוסיפו טיפה של מדיום הרכבה לכל באר, והימנעו מבועות. הניחו מכסה על גבי בארות הזרעים. כסו במגבוני נייר ולחצו בחוזקה על הכיסוי כדי להסיר את אמצעי ההרכבה העודפים. היזהרו לא להזיז את הכיסוי, ואל תנקו את אמצעי ההרכבה העודפים. צלם מספר תמונות עם מיקרוסקופ הפוך עם אור שדה בהיר והגדלה מינימלית של 40x. בצע בדיקת multiome.המשך מיד אל Chromium Next GEM Single Cell Multiome ATAC + Gene Expression מדריך למשתמש (CG000338 – Rev F)25. 2. טיהור גרעינים מתאי גזע ואב המטופויטיים של מח עצם עכבר (HSPCs) הערה פרוטוקול זה מתאר את טיהור הגרעינים משלוש תת-קבוצות של מח עצם HSPCs: שושלת-c-Kit+Sca-1+ תאי גזע המטופויטיים (HSC), שושלת-c-Kit+Sca-1-CD34+FcγR- אבות מיאלואידים נפוצים (CMP), ושושלת- c-Kit+Sca-1-CD34+FcγR+ אבות גרנולוציטים-מונוציטים (GMP). יש ללבוש כפפות לטקס או ניטריל בכל עת במהלך ההליך. פרוטוקול זה הוא אדפטציה של פרוטוקול 10X Genomics Demonstrated – בידוד גרעינים עבור Multiome ATAC + GEX sequencing (CG000365 – Rev C)27. שינויים הוכנסו בפרוטוקול המקורי כדי למקסם את התאוששות הגרעינים. מומלץ מאוד שיהיו שני אנשים שיבצעו את הניסוי, שיהיו להם שלבים 1. ל-3. (כלומר, הכנת התמיסה החד-תאית) המבוצעת על ידי אדם אחד, ושלב 4 (כלומר, הכנת המיון) המבוצע במקביל על ידי אדם אחר. מכיוון שהפרוטוקול רגיש מאוד לזמן, קריטי למזער את זמן עיבוד הדגימה על ידי הכנת הממיין מיד עם הכנת המתלה החד-תאי. הכנת ריאגנטים וחומריםממלאים דלי בקרח. הכן את מאגר FACS: DPBS עם פתרון HI-FBS 2% (כ-500 מ”ל עבור 6 דגימות) וסנן דרך מסנן 0.2 מיקרומטר. הכן את אמצעי האיסוף: DPBS עם תמיסת 10% HI-FBS (500 מיקרוליטר לדגימה), וסנן דרך מסנן 0.2 מיקרומטר. בידוד תאי מח עצםלהקריב עכברים באמצעות פרוטוקולים שאושרו על ידי המוסד. בניסוי זה, העכברים הוקרבו על ידי פריקת צוואר הרחם לאחר מנת יתר של קטמין/קסילזין. רססו את הבטן והרגליים האחוריות של העכברים באתנול 70%. השתמשו במלקחיים סטריליים ובמספריים כדי לבצע חתך קטן באמצע הבטן התחתונה ולפתוח את הצפק מבסיס הרגליים האחוריות לסרעפת (איור משלים 1). בצעו חתך נוסף עבור כל רגל אחורית בניצב לצפק הפתוח, ואז תפסו את אחד הצדדים של אחד החתכים הנוספים האלה ומשכו אותו בנפרד כדי לקלף את העור משתי הרגליים האחוריות מעבר למפרק הקרסול כדי לחשוף את השרירים של שתי הרגליים האחוריות (איור משלים 1A). רפדו את המספריים לאורך עמוד השדרה במפרק הירך של רגל אחורית אחת כדי לחתוך את הרגל מבלי לחתוך דרך עצם הירך (איור משלים 1B, C). חזור על אותו הדבר עבור הרגל השנייה. כדי לבודד את עצם הירך, חתכו את רוב רקמת השריר החוצה, ואז החזיקו את עצם הירך ואת השוקה בכל יד עם קצות האצבעות במפרק (איור משלים 1D, E). קפלו בעדינות את הרגל כנגד העיקול הטבעי כדי לנתק את השוקה מעצם הירך (איור משלים 1E) ולאחר מכן חתכו בזהירות את רקמת החיבור עם מספריים כדי להפריד בין עצם הירך לשוקה. השתמשו במספריים בתנועות פיתול קלות כדי להזיז את פיסת עמוד השדרה מהקצה העליון של עצם הירך (איור משלים 1E). נקו את עצם הירך המבודדת עם נייר טישו כדי להסיר את השריר ורקמת החיבור שנותרו. יש לשמור קר על קרח בצלחת בת 12 בארות מלאה היטב ב-2 מ”ל DMEM (1x) + GlutaMAX-I. לאחר שכל עצם הירך נאספת, ודא כי רקמות שריר וסיביות מוסרים לחלוטין מן העצם. אין לחתוך את העצם פתוחה כדי (א) לשמור על המח בתוך סטרילי ו-(ב) להימנע מאיבוד תאים בבאר. השתמש בשלבים הבאים לשטיפת התאים משתי עצמות הירך של עכבר אחד, המותאמות מהאג ומורטי28. הכינו צינור אחד של 1.5 מ”ל וצינור אחד של 0.5 מ”ל. הוסף 150 μL של חיץ FACS לצינור 1.5 מ”ל, ולאחר מכן תקע חור בתחתית הצינור של 0.5 מ”ל באמצעות מחט 18 G והכנס את הצינור 0.5 מ”ל לצינור 1.5 מ”ל. פתחו את החלק הדיסטלי של כל עצם הירך באמצעות מספריים כירורגיים של עכברים (איור משלים 1F): נעלו את האפיפיזה הדיסטלית בין הלהבים והפעילו לחץ עדין תוך כדי היפוך המספריים כדי לנתק בצורה חלקה את האפיפיזה הדיסטלית מבלי לפתוח את העצם בחוזקה. אם זה מצליח, 4 בליטות אמורות להיראות בקצה הפיזי החשוף כעת (איור משלים 1G). התאימו את שתי עצם הירך כשהקצה הפתוח פונה כלפי מטה לתוך צינור 0.5 מ”ל מוכן הממוקם בתוך צינור של 1.5 מ”ל המכיל חיץ FACS (איור משלים 1H). הניחו מסננת תאים של 70 מיקרומטר על צינור של 50 מ”ל והרטיבו מראש את המסננת עם 2 מ”ל של חיץ FACS. כדי לשטוף את מח העצם, צנטריפוגה את הצינורות (כובעים פתוחים) ב 12,000 x גרם עד הצנטריפוגה מגיע לערך 12,000 x גרם , ואז מיד לעצור את הצנטריפוגה. ודאו שתאי מח העצם מכווננים בצינור של 1.5 מ”ל ושעצם הירך לבנה (לפני שטיפת התאים היא אדומה) (איור משלים 1I). יש להשליך את צינורות ה-0.5 מ”ל עם 2 עצם הירך. השליכו את הסופרנאטנט 150 μL באמצעות פיפטה. יש להשהות מחדש את הגלולה עם מיקרופיפטה ב-1 מ”ל של חיץ שוכב אמוניום-כלוריד-אשלגן (ACK) למשך 1-2 דקות ב-RT כדי לליז תאי דם אדומים. הימנעו מזמני דגירה ארוכים יותר מכיוון שהם עלולים לגרום לירידה בכדאיות של תאים בעלי גרעין. מעבירים לצינור 50 מ”ל דרך מסננת התאים 70 מיקרומטר שהורטבה מראש. הוסף 10 מ”ל של חיץ FACS כדי לדלל את מאגר ACK שוכב ובכך להפסיק את הליזיס. צנטריפוגה ב 400 x גרם במשך 5 דקות ב 4 ° C. השהה מחדש במאגר FACS של 10 מ”ל על ידי השעיה מחדש תחילה ב- 1 מ”ל ולאחר מכן מילוי ב- 9 מ”ל. הכן את התאים לספירה כמתואר ב- 1.3.8. ספור את התאים באמצעות מונה תאים אוטומטי בהתאם להמלצות הספק. הוא צפוי לאסוף כ-40 מיליון תאים מ-2 עצם הירך. צביעת מח עצם HSPCצנטריפוגה את התאים ב 400 x גרם במשך 5 דקות ב 4 ° C ו resuspend את הגלולה עם micropipette במאגר FACS לריכוז סופי של 1 x 107 תאים / מ”ל. עם מיקרופיפטה P1000, העבר את המתלה לתוך צינור FACS, סינון דרך מכסה מסננת תאים 35 מיקרומטר. הכינו דגימות מבחנה של כתמים בודדים עבור כל נוגדן המופיע בטבלה 1 כדי להגדיר פיצויים של פלואורוכרום על ממיין התאים:הכינו צינור FACS אחד לכל נוגדן ומלאו את הצינורות ב-200 מיקרוליטר של PBS. הוסף 15 μL של חרוזי פיצוי פלואורוכרום בכל צינור FACS של נוגדן מצומד פלואורוכרום. בצינורות FACS עבור תאים מוכתמים לא מוכתמים ועבור תאים מוכתמים בודדים חיים/מתים, הוסף 500,000 תאים במקום חרוזים. הוסף 1 μL של כל נוגדן מצומד פלואורוכרום (ראה טבלה 1) לתוך צינור FACS המתאים לו. הוסף 0.5 μL של כתם חי/מת לצינור FACS של כתם יחיד חי/מת. יש לשמור על קרח למשך 15 דקות כשהוא מוגן מאור. הכינו תערובות 1 ו-2 כמצוין בטבלה 2.הערה: נפחי הנוגדנים המצוינים בטבלה 2 תקפים עבור הנוגדנים המוזכרים בטבלת החומרים. הם צריכים להיות מותאמים לכל התייחסות נוגדנים חדשה או הרבה שונה של אותה התייחסות נוגדנים. הוסיפו 300 μL של Mix 1 לצינור הדגימה, השהו מחדש ושמרו למשך 15 דקות על קרח מוגן מפני אור. הוסיפו 300 μL של Mix 2 לצינור הדגימה, השהו מחדש ושמרו למשך 20 דקות על קרח מוגן מפני אור. הוסף 3 מ”ל של מאגר FACS לצינורות המוכתמים הבודדים ולצינורות הדגימה המוכתמים במיקס. יש לסחור כלפי מטה במהירות של 400 x גרם למשך 5 דקות ב-4°C. בזהירות להשליך את supernatant באמצעות micropipette ו resuspend את הגלולה ב 500 μL של מאגר FACS. הכינו צינור של 1.5 מ”ל ממולא מראש ב-500 מיקרוליטר של אמצעי איסוף.הערה: מיקס 1 מוכן ב-DPBS מכיוון שהוא מכיל את הכתם חי/מת המושפע באופן משמעותי מ-HI-FBS. לאחר שהתאים מוכתמים על ידי Live/Dead, מתווסף Mix 2, המכיל נוגדנים מצומדים פלואורוכרום התלויים מחדש במאגר FACS המכיל HI-FBS. היוצא מן הכלל היחיד הוא נוגדן אנטי CD16/32 הכלול ב- Antibody Mix 1 כדי לשמש כחוסם קולטן Fc המונע קשירה לא ספציפית של נוגדנים אחרים שנוספו בשלב הבא. מיון תאים באמצעות FACSהערה: בעוד שניתן לבצע מיון תאים במגוון רחב של ממייני תאים, כאן מתואר הליך השימוש במכשירי BD FACSAria Fusion או BD FACSAria III. מומלץ מאוד לבצע את הכיול וההתקנה של ממיין התאים תחת פיקוח או על ידי משתמש מנוסה במכשיר.כיול מכשיר FACS: עיין בפרוטוקול 1 שלב 4.1. הגדרת מכשיר FACS למיון תאים:התחל את רכישת התאים המוכתמים. אלה משמשים להגדרת הפיזורים קדימה וצידית ואת מתח הגלאי עבור כל פלואורופור. הגדר את הפרמטרים כך שהאות הפלואורסצנטי של כל פלואורופור ייפול בתוך העשור הראשון של סולם היומן בתרשים הנקודות. רכוש פקדי צבע בודדים כדי להגדיר פיצויים באופן ידני (יש ליישר חציון של אוכלוסיות חיוביות ושליליות) או השתמש בתוכנת החישוב האוטומטית (מדידות שיפוע). ודא שבקרות הפיצוי תואמות את פלואורוכרומי הניסוי ואת הגדרות הגלאים. הקלט 10,000 אירועים עבור תאים ו- 5,000 אירועים עבור חרוזים. השתמשו בצינורית הדגימה (כלומר, תאים מרובי מוכתמים) כדי להגדיר אוכלוסיות תאים מעניינות באמצעות אסטרטגיית ה-gating שמוצגת באיור 3A. בצע את שלבים 4 – 6 (להלן). כדי לזהות את שלושת מח העצם HSPCs של עניין (HSC, CMP, ו GMP), להתחיל את gating באמצעות גודל (FSC-A) וגרעיניות (SSC-A) כדי שער על לויקוציטים, ולאחר מכן FSC-H/FSC-A כדי להפלות כפולים. מבוסס על סמן SSC-A/תא מת, שער תאים חיים. השתמש ב- Lineage/c-Kit כדי לבחור תאים שליליים לשושלת ומבטאים רמות ביניים עד גבוהות של c-Kit. באמצעות c-Kit/Sca-1, שער על שושלת c-Kit+ Sca-1+ (LKS+) HSCs, אחת משלוש האוכלוסיות המעניינות. בין האבות המיאלואידים (שושלת-c-Kit+Sca-1-), השתמש ב- FcγR/CD34 כדי לא לכלול CD34-FcγR- מגה-קריוציטים ואבות אריתרואידים (MEP), תוך הכללת CD34+ FcγR- CMP, כמו גם CD34+FcγR+ GMP באוכלוסיות התאים שיש למיין. ודא שהזרם והסטייה יציבים. במצלמת הזרם הצדדי, הפעל את מיון הבדיקה, הפעל מתח ואשר את מיון הטיפות המדויק בצינור 1.5 מ”ל המותקן בצד שמאל. בחלון פריסת מיון , בחר את האוכלוסיות המעניינות (כלומר, “LKS+” ו- “CD34+ אבות מיאלואידים” המוצגים בדוגמה זו). תחת התקן, בחר 2 צינור. תחת דיוק, בחר טוהר. באירועי יעד, בחר רציף כדי למיין בין 160,000 ל- 200,000 אבות מיאלואידים LKS+ ו- CD34+ . הוסף 500 μL של מאגר FACS לתרחיף התא והעבר את 1 מ”ל הדגימה על ידי סינון לתוך צינור FACS חדש של 35 μm מסננת תאים כדי להבטיח שכל התאים נמצאים בתרחיף יחיד ממש לפני הרכישה. זה מבטל גושי תאים שעלולים לסתום את המכשיר. לאחר שתהיה מוכן, לחץ על מיין ואישור כדי להתחיל במיון. התאם את קצב הזרימה כדי לשמור על מהירות מתחת ל-10,000 אירועים בשנייה.הערה: היחס הצפוי בין אבות מיאלואידים LKS+ ל- CD34+ הוא 1:3 עבור עכבר נקבה C57BL/6J בוגר (בן 8-12 שבועות) במצב יציב. מספרי התאים הממוינים הממוקדים מגיעים בדרך כלל תוך 30 דקות מהמיון. בקרת איכות וספירה של תאים ממויניםהערה: שלב זה ייעשה רק במהלך ניסוי הפיילוט לאופטימיזציה של שלבי הכנת הדגימה, במטרה לבדוק את טוהר התאים הממוינים שישמשו לבידוד גרעינים. לאחר אופטימיזציה מלאה של הפרוטוקול, לא מומלץ לבצע שלב בקרת איכות זה בניסויי המעקב כדי למנוע בזבוז מיותר של חומר מוצא שעשוי להיות זמין במספרים נמוכים לבידוד גרעינים.בקרת טוהר על ידי ציטומטריית זרימההעבר 10 μL של התאים הממוינים לתוך צינור FACS חדש המכיל 90 μL של מאגר FACS. רכוש ותעד נתונים לאחר המיון כדי לאמת את טוהר המיון ואת הכדאיות. ודא שלפחות 95% מהתאים מופיעים בשער העניין, כפי שהוגדר ב-3-6 וכפי שמודגם באיור 3B. בידוד גרעינים ממח עצם ממוין HSPCsהשתמש בפרוטוקול “בידוד גרעיני קלט תאים נמוך” של הנספח מתוך פרוטוקול 10X Genomics Proven Protocol – בידוד גרעינים עבור ריצוף ATAC + GEX Multiome של תא בודד (CG000365 – Rev C)27, עם השינויים הבאים שבוצעו כדי לייעל את התאוששות הגרעינים:זמן ליזיס: הפעל ניסוי פיילוט עבור פרוטוקול זה כדי לזהות את זמן הליזיס הטוב ביותר לבידוד גרעינים. הקפד להשיג ליזה שלמה של התא תוך שמירה על גרעינים שלמים.הערה: שלב f של פרוטוקול 10X Genomics27 שהוזכר לעיל מורה “לדגור [במאגר Lysis] במשך 3-5 דקות על קרח”. במהלך ניסוי הפיילוט, בדקו לפחות 3 דקות, 4 דקות ו-5 דקות והעריכו את כמות הגרעינים המשוחזרים על ידי ספירה ואיכות על ידי ציטומטריית זרימה והדמיית מיקרוסקופיה כדי לבחור את משך הליזה האופטימלי (ראו תיאור של בדיקות בקרת איכות אלה להלן). כדי לחסוך ריאגנטים, החלף את מאגר הגרעינים המדולל ב- PBS 0.04% BSA בניסוי הפיילוט. עבור מח עצם HSPCs, 3 דקות זוהו כמשך הליזה האופטימלי. צנטריפוגות תאים: עבור כל הצנטריפוגות התלויות תאים, צנטריפוגה ב 300 x גרם במשך 7 דקות (במקום 5 דקות CG000365 – Rev C)27 ב 4 ° C. צנטריפוגות גרעינים: בצע את כל צנטריפוגות תרחיף הגרעינים ב 500 x גרם במשך 5 דקות לפי CG000365 – Rev C27. איסוף גרעינים: בשלב b, לאחר השעיה מחדש של 50 μL של PBS 0.04% BSA והעברה לצינור 0.2 מ”ל, הוסף 50 μL של PBS 0.04% BSA לצינור המקורי ותערובת פיפטה כדי לאסוף את שאריות התאים. מעבירים לצינור 0.2 מ”ל כדי להגיע לנפח כולל של 100 מיקרוליטר. מעתה, הנפח הכולל יהיה 100 μL במקום 50 μL של הפרוטוקול. התאם את השלבים במורד הזרם בהתאם (לדוגמה, עבור שלב d, הסר 90 μL במקום 45 μL; עבור שלב e, הוסף 90 μL של חיץ ליזיס במקום 45 μL). עבור שלב m, השהה מחדש את גלולת הגרעינים ב- 12 μL של מאגר גרעינים מדוללים במקום 7 μL. ספרו את הגרעינים המבודדים. לתוך צינור ריק 0.5 מ”ל, להוסיף 10 μL של 0.4% כחול טריפאן ו 8 μL של PBS 0.04% BSA. הוסף 2 μL של גרעינים לצינור וספור גרעינים כמתואר ב- 1.3.8. השתמש במונה תאים אוטומטי בהתאם להמלצות הספק. בקרת טוהר על ידי ציטומטריית זרימההערה: שלב זה ייעשה רק במהלך ניסוי הפיילוט לאופטימיזציה של שלבי הכנת הדגימה לבדיקת טוהר הגרעינים שיש לטעון על שבב 10X Chromium. לאחר אופטימיזציה מלאה של הפרוטוקול, לא מומלץ לבצע שלב בקרת איכות זה בניסויי המעקב כדי למנוע בזבוז מיותר של גרעינים שנאספו שעשויים להיות זמינים במספרים נמוכים.לאחר השלמת בידוד גרעינים, העבירו 6 μL של תרחיף גרעינים בצינור FACS חדש שמולא מראש ב-150 μL של מאגר FACS. מוסיפים 3 μL של 7-AAD ודגרים במשך 5 דקות על קרח. רכוש ותעד נתונים לאחר המיון כדי לאמת את טוהר המיון ואת הכדאיות. יש לוודא שלפחות 95% מהגרעינים מופיעים בשער העניין, כהגדרתו בפרוטוקול 1 שלב 4.2 (ראה איור 4). בקרת איכות של גרעינים מטוהרים במיקרוסקופיה:הערה: שלב זה ייעשה רק במהלך ניסוי הפיילוט לצורך אופטימיזציה של שלבי הכנת הדגימה לבדיקת איכות הגרעינים שיש לטעון על שבב 10X Chromium. לאחר אופטימיזציה מלאה של הפרוטוקול, לא מומלץ לבצע שלב בקרת איכות זה בניסויי המעקב כדי למנוע בזבוז מיותר של גרעינים שנאספו שעשויים להיות זמינים במספרים נמוכים.המשך כמתואר בשלב 1.5.3. ביצוע בדיקת מולטיוםהמשך מיד אל Chromium Next GEM Single Cell Multiome ATAC + Gene Expression מדריך למשתמש (CG000338 – Rev F)25.