The immobilization device is a bilayer PDMS block fabricated by bonding two layers: a flow layer (Layer 1) and a control layer (Layer 2) as shown in Figure 1. The main trap is connected to a nitrogen gas cylinder through a regulator and a 3-way stop cock to apply necessary (3-14 psi) pressure onto the membrane through a liquid column (Figure 1A). The deflected membrane immobilizes C. elegans, Drosophila or zebrafish larvae in the flow channel designed with different dimensions (Figure 1B and 1C). Photo masks are used to design immobilization device layers with different geometries for C. elegans (Figure 1D), Drosophila larvae (Figure 1F) and zebrafish larvae (Figure 1G). The channel height of the flow layer (h1 in Figure 1E, 1K) was varied using different spin coating speeds to accommodate organisms of different diameters inside the flow channel. Flow layer heights were fabricated at 40 μm, 80 μm, 500 μm and 900 μm for C. elegans larval stages, adult C. elegans, first instar Drosophila larvae and zebrafish larvae respectively (Table 1). The device with a flow channel height of 40 μm is appropriate from L1 to early L4 C. elegans larvae. A device with an 80 μm flow channel height is used for late L4 larvae and adult C. elegans when the vulva starts to protrude. The larger zebrafish devices can also be used for older Drosophila larvae. The PDMS membrane thickness (m) was fabricated at 40 μm and 300 μm for C. elegans and Drosophila/zebrafish devices respectively. The height of the microfluidic channel in layer 2 was fabricated at 40 μm for early larval stages of C. elegans and 80 μm for late larval stages of C. elegans and Drosophila/zebrafish larvae. The height of the flow channel, membrane and control channel were measured in devices fabricated in multiple independent batches and were found to be consistent within ± 5 μm of each other. The device uses a membrane deflection technique (Figure 1H-J) to immobilize organisms in the flow channel.
| Organisms | Flow channel (PDMS-1) | Spin conditions for flow channel | Spacer PDMS (PDMS-S) | Spin conditions for spacer PDMS | Immobilization pressure (psi) |
| C. elegans (L1 to early L4) | 40 μm (SU8-2025) | 500 rpm (5 sec), 2,000 rpm (30 sec) | NA | NA | 14 |
| C. elegans (late L4 and adults) | 80 μm (SU8-2050) | 500 rpm (5 sec), 2,000 rpm (30 sec) | NA | NA | 14 |
| Drosophila (first instar) | 80 μm (SU8-2050) | 500 rpm (5 sec), 2,000 rpm (30 sec) | 400 μm (PDMS) | 500 rpm (35 sec) | 7 |
| Zebrafish (30 hpf) | 80 μm (SU8-2050) | 500 rpm (5 sec), 2,000 rpm (30 sec) | 2X 400 μm (PDMS) | 500 rpm (35 sec) | 3 |
Table 1. Device dimensions of the flow channel (PDMS-1) and spacer PDMS layer (PDMS-S) used in C. elegans and Drosophila/zebrafish devices.
C. elegans L4 larvae were immobilized in the 80 μm channel height PDMS microfluidic device using 14 psi nitrogen gas (Figure 2A). Time-lapse fluorescence images were acquired at the speed of 2 frames per second (fps) using a 60X objective (numerical aperture 1.4, oil objective) for imaging transport of mitochondria visualized using a mitochondrial matrix targeted GFP in the touch receptor neurons 21. All 400 frames were converted to kymographs using ImageJ (www.rsbweb.nih.gov/ij) and mitochondria were classified as anterogradely directed (away from the cell body), retrogradely directed (towards the cell body) or stationary (Figure 2E). We observed mitochondrial transport up to 21 psi of immobilization pressure, but the flux was somewhat greater at the lowest immobilization pressure. The anterograde and retrograde flux of mitochondria were measured to be 1.1 ± 0.23 and 1.6 ± 0.22 at 7 psi whereas 0.8 ± 0.25 and 1.2 ± 0.34 at 14 psi immobilization pressure. The values were not statistically different from 1.1 ± 0.33 and 1.3 ± 0.42 obtained from animals immobilized using 3 mM levamisole (p-values>0.45, n=30 animals).
The larger devices with 500 μm height can be used to immobilize first instar Drosophila larvae (Figure 2B) and 28-30 hpf zebrafish larvae (Figure 2D). Dissection of first instars Drosophila larvae can be challenging. By contrast microfluidic devices provide a method to perform high resolution bright field and/or fluorescence imaging using intact organisms. Individual Drosophila larvae were immobilized using 7 psi of compressed nitrogen gas (Figure 2B, Table 1) and fluorescence images of synaptotagmin.eGFP transport in sensory neurons were acquired (Figure 2C). Time-lapse movies showed moving and stationary synaptotagmin marked cargo in sensory neurons (Figure 2F). Average anterograde and retrograde velocity was measured from kymographs to be 0.92 ± 0.04 μm/s and 1.00 ± 0.05 μm/s respectively (n=6 animals, n>40 segments). These are comparable to velocities of synaptotagmin carrying transport vesicles measured in dissected Drosophila motor neurons moving both anterogradely (0.84 ± 0.05 μm/s ) and retrogradely (0.76 ± 0.03 μm/s) 22.
Similar devices with 900 μm height were used to immobilize different stages of zebrafish larvae (Figure 2D, Table 1). Zebrafish larvae during its early developmental stages flips its tail every 10 to 15 sec and are usually immobilized using 0.02 % tricaine (MS-222) in solution or on mounting media for high resolution imaging 23. Our PDMS device provides an alternate means for fast immobilization and eliminates the need of an unreliable mounting medium in the optical path during high resolution imaging 24. We manually dechorionated 28-30 hpf larvae and immobilized them individually in a PDMS device using 3 psi nitrogen gas to acquire time-lapse movies of larval heartbeat. The rate of heartbeat was measured to be 136.8 ± 1.6 per min (n=8 movies) and was similar to other published reports 25.
Our simple microfluidic device had already been demonstrated for measuring a variety of sub-cellular and cellular events in wild type C. elegans 4. In our microfluidic device, wild type C. elegans show a larger number of cargo such as synaptic vesicles that move in neurons compared to animals that are immobilized using anesthetics. However, we had not tested our devices for mutant animals to see if these findings hold true. To test this we used a strong hypomorphic mutant in the kinesin3/UNC-104 molecular motor, unc-104(e1265), that transports pre-synaptic vesicles 26. We compared the flux of GFP::RAB-3 marked pre-synaptic vesicles in anterior lateral mechanosensory neurons in anesthetic immobilized and device immobilized mutant aniamls (Figure 3A). Flux of pre-synaptic vesicles was greater flux in device immobilized animals as compared to data acquired from unc-104 animals anesthetized in 1 mM levamisole (Figure 3B, 3C) 27. This shows that imaging strong transport defective mutants in microfluidic devices is likely to be a more efficient means of collecting data. We also show that other cargo e.g. dendritic transport of glutamate receptors in URY neurons can be imaged in device immobilized animals (Figure 3D, 3E). The device can also be used to image cellular processes during early C. elegans development such as Q neuroblast division in L1 animals. The QR cell was imaged to divide to form two daughter cells QR.a and QR.p ~3 hr after hatching (Figure 3F) consistent with earlier observations 28.
Table 1. Device dimensions of the flow channel (PDMS-1) and spacer PDMS layer (PDMS-S) used in C. elegans and Drosophila/zebrafish devices.
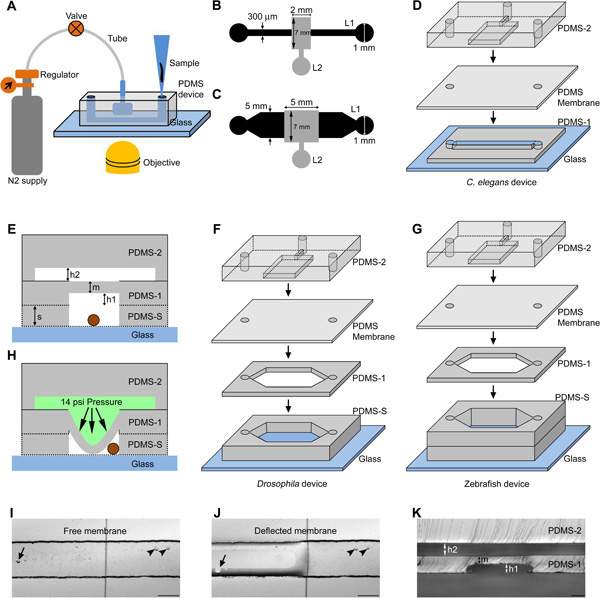
Figure 1. Schematic representation of PDMS microfluidic devices for genetic model organisms. (A) The organism is loaded using a micro tip into the flow channel and immobilized using compressed nitrogen gas controlled using a regulator and valve. The device is suitable for bright field/fluorescence imaging in an inverted microscope. The design consists of a 10 mm long flow channel (design I, L1) and a control channel (design II, L2) respectively for C. elegans (B) and Drosophila/zebrafish larvae (C). The dimensions are indicated on the design (B and C). (D, F, G) Schematic of various layers of PDMS (PDMS-1, PDMS-2, PDMS membrane, PDMS-S and Glass) present in a C. elegans, Drosophila and zebrafish immobilization device. (E) Schematic representation of a cross section of the device showing the bulk PDMS layer (PDMS-2), a spin coated PDMS layer (PDMS-1) to form a flexible membrane and a spacer PDMS layer (PDMS-S) to add height to the flow channel (for Drosophila/zebrafish device). The whole structure is bonded irreversibly to an optical grade glass cover slip to accommodate the organism (circle in brown). The schematic indicates height of the control channel ‘h2’, flow channel ‘h1’, punched spacer layer ‘s’ and membrane thickness ‘m’. (H) Indicates the membrane deflection towards the flow channel in the presence of liquid column under pressure in the control channel. Bright field images are shown of a free (I) and deflected membrane (J) under zero and 14 psi pressurized liquid column respectively. The arrow and arrow head indicate bubble under and out of the membrane in the flow channel respectively. (K) Cross sectional image of the C. elegans device taken at the location of the immobilization area. Scale bar is 50 μm (K) and 200 μm (I, J). (D, F, G) Schematics are not to scale. Click here to view larger figure.
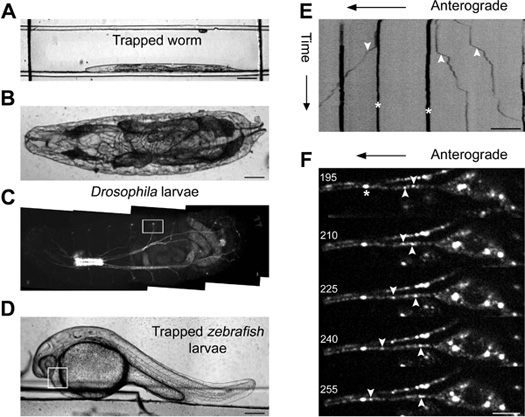
Figure 2. Imaging genetic model organisms immobilized in a microfluidic device. Bright field image of an immobilized C. elegans (A), a first instar Drosophila larvae (B) and a 30 hpf zebrafish larvae (D) in a PDMS microfluidic device. Kymograph analysis of time-lapse imaging of jsIs609, a C. elegans strain, expressing a mitochondrial matrix targeted GFP in an anterior lateral mechanosensory neuron (ALM) of a device immobilized animal (E). Mitochondria are classified as anterograde (‘down’ arrow head), retrograde (‘up’ arrow head) and stationary (‘star’ mark) in the kymograph. (C) Fluorescence image of an intact first instar Drosophila larva. The box indicates the location where high resolution time-lapse imaging of syt.eGFP transport is carried out in a cholinergic neuron. Montage of five frames acquired at 5 Hz with frame numbers indicated on each image and GFP marked cargo shown as anterograde, retrograde and stationary in the montage (F). The box in (D) indicates the area that was monitored for calculating rate of heartbeat of zebrafish larvae. Scale bar is 5 μm (F), 10 μm (E), 100 μm (A, B) and 200 μm (D).
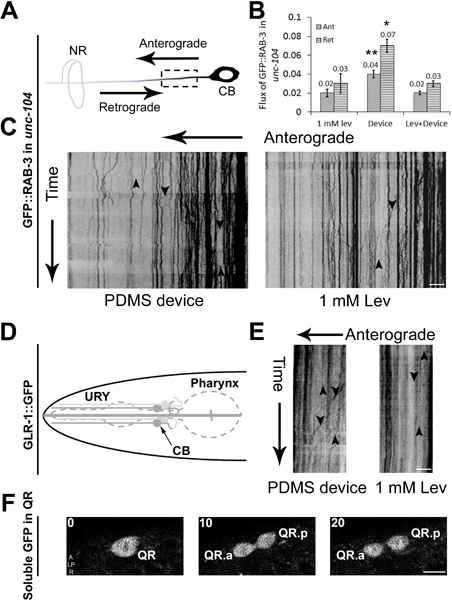
Figure 3. Cellular and sub-cellular imaging in C. elegans using PDMS microfluidic devices. (A) Schematic of an anterior lateral mechanosensory (ALM) neuron with the anterograde and retrograde directions marked. The dotted box shows the region imaged for axonal transport. 350 frames are acquired using a spinning disc confocal microscope at 5 fps with cell body (CB) on the right and nerve ring (NR) on the left. Data are analyzed using kymographs for animals immobilized in a device or using 1 mM levamisole (C). Cargo moving in anterograde and retrograde directions are shown using arrows pointing ‘down’ and ‘up’ respectively. (B) Anterograde and retrograde flux of particles observed in unc-104(e1265) animals in a 20 μm region of the ALM neuron over 350 time-lapse frames. (D) Schematic of an URY neuron used to image GFP marked glutamate receptors transport. 350 time-lapse frames are converted into kymographs that show cargo moving in the anterograde and retrograde directions in animals immobilized in PDMS device or using 1 mM levamisole (E). Time-lapse images acquired during Q neuroblast division and migration in early larval stages of C. elegans. (F) Three time-lapse frames that show QR undergoing division to form its daughter cells QR.a and QR.p. Scale bar is 5 μm (C, E and F). Data represented in (B) is mean ± SEM (n>4). Comparisons are made with respect to values obtained in anesthetic and denoted by * (p<0.05) and ** (p<0.005).
