Adult rat spinal cord cells grown in suspension culture in EFH medium will form small neurospheres (colonies of undifferentiated cells) within 1 week of initial plating. In primary cultures, most of the cells plated will die and growth factor-responsive stem cells will proliferate and are selected for in the EFH medium. By passage 3, there will be numerous free-floating neurospheres about 100 µm in diameter (Figure 2A). Neurospheres are round and phase-bright, and under high magnification, cilia-like microspikes are seen protruding from cells on the outside of spheres which are characteristic of neurospheres unlike clumps of cell debris (Figure 2B). The typical yield of cells from rat tissue is about 2 x 106 cells with about 80% viability. The neurospheres should not be seeded at too high a density to avoid aggregation of cell clusters. Also, large neurospheres become difficult to dissociate and cells will become necrotic in the center of the sphere. This will also occur if cultures are left too long before passaging. In EFH culture, adult rat spinal cord NSPCs proliferate (Figure 2C) and express nestin (Figure 2D) a marker for precursor cells, and low levels of mature neural markers (data not shown).
Adult human spinal cord NSPC cultures do not grow as rapidly as rat NSPC cultures. If human spinal cord cells are initially cultured in suspension, they will form aggregates and irregular clusters of small numbers of cells combined with debris (Figure 3A). Subsequent passaging of these cultures does not promote enrichment of the NSPC population. However, when the human cells are plated in EFH on an adherent monolayer, the growth factor-responsive NSPCs adhere to the substrate (Figure 3B) and subsequent media replacement removes myelin and cell debris (Figure 3C). The typical yield of cells from human tissue is about 1 x 106 cells with about 70% viability. When the human NSPC cultures become well established as an adherent monolayer, the NSPCs can then also be plated in suspension cultures to form free-floating neurospheres. In EFH culture, adult human spinal cord NSPCs proliferate (Figure 3D) and express nestin (Figure 3E) and Sox2 (Figure 3F), a transcription factor shown to be expressed by neural stem cells, and very low levels of mature neural markers (data not shown).
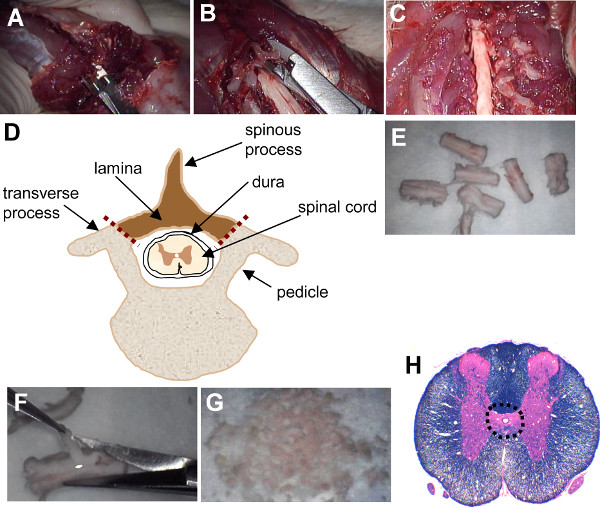
Figure 1. Laminectomy of rat spinal cord and dissection of periventricular region. (A) At the exposed caudal end of the spinal cord, rongeurs are inserted extradurally into the lateral aspect of the spinal canal. (B) Small cuts are made into the lamina on each side of the vertebrae and the lamina is carefully peeled away to expose the spinal cord. (C) The exposed cervical spinal cord after laminectomy. (D) Schematic of thoracic vertebral cross-section showing spinal cord and location of cuts made for laminectomy (depicted with red dotted lines). The laminae and spinous process (shaded region) are removed. (E) The spinal cord is cut transversely into 1 cm segments. (F) The overlying meninges, white matter, and most of the grey matter is carefully removed using microscissors. (G) Minced periventricular tissue. (H) Transverse section of rat spinal cord stained with luxol fast blue and hemotoxylin and eosin with dotted outline showing dissected periventricular region. Please click here to view a larger version of this figure.
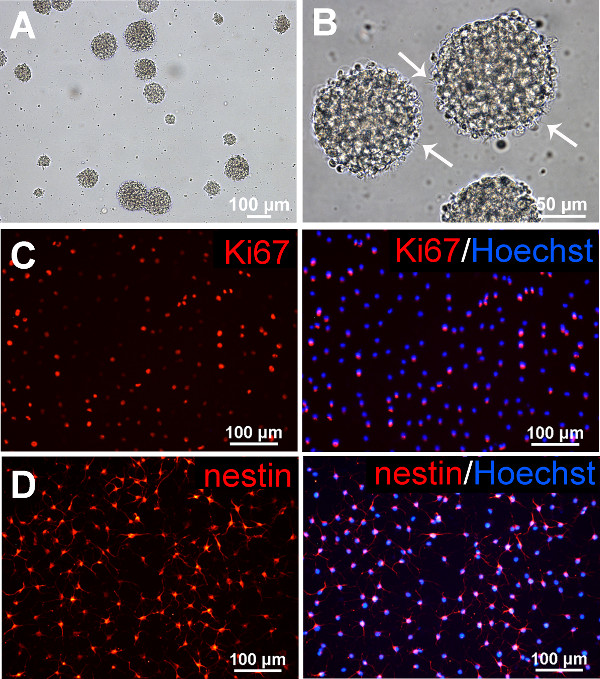
Figure 2. Adult rat spinal cord NSPCs. (A) By passage 3, numerous neurospheres are seen when rat spinal cord NSPCs are grown in free-floating suspension culture. (B) Neurospheres are phase-bright and microspikes protruding from the neurospheres (arrows) are apparent at high magnification. (C) Dissociated neurospheres (passage 3, day 4) are plated on Matrigel-coated wells in EFH medium, fixed, and immunostained and counterstained with Hoechst to visualize nuclei (blue). In EFH conditions, adult rat spinal cord NSPCs proliferate (as shown with Ki67 immunostaining), and (D) primarily express nestin. Please click here to view a larger version of this figure.
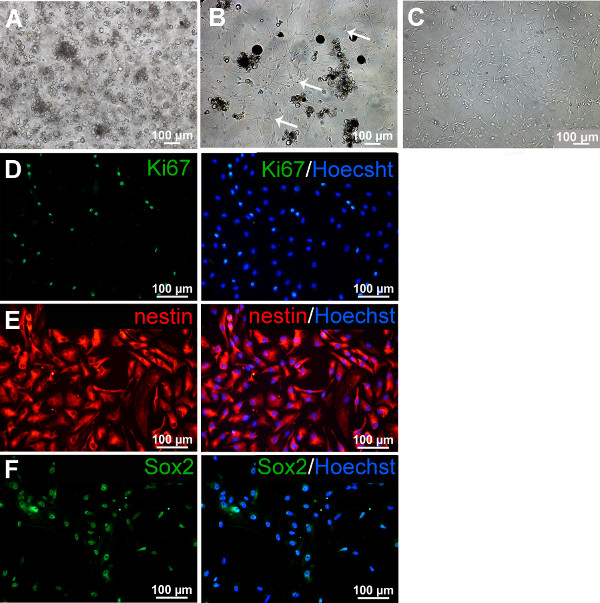
Figure 3. Adult human spinal cord NSPCs. (A) When initially plated in culture flasks, adult human spinal cord NSPCs do not grow well. Primary free-floating cultures in EFH medium (13 days after plating) show aggregates of cells and debris. (B) In contrast, in primary adherent cultures in EFH, NSPCs have attached to the Matrigel substrate (arrows), shown at 17 days after plating, and (C) at 41 days in vitro. (D) In EFH medium, adult human spinal cord NSPCs proliferate, as shown with Ki67 immunostaining (26 days in vitro shown), and primarily express (E) nestin (26 days in vitro shown) and (F) Sox2 (66 days in vitro shown). Nuclei are counterstained with Hoechst (blue). Please click here to view a larger version of this figure.
