Специализированный методы напряжение зажимные разрешить запись ионных токов при контролируемых мембранных потенциалов. Широко используется двухэлектродной зажим напряжения (TEVC) и методы патч зажим обеспечивают надежную электрофизиологические информацию о свойствах многих ионных каналов. Тем не менее, оба этих метода имеют свои недостатки, которые препятствуют приобретению надежных данных для быстрых напряжения натриевых каналов и других каналов быстро активирующих в мембранах, таких как те, из Xenopus ооциты. В Безания и Стефани лаборатории следовательно разработана методика Вазелин разрыв зажим напряжения бортового открытым (CoVG) для ооцитов 2. Методика была широко применяться для записи, Na +, K + и Ca 2 + каналов 3-8.
Во время записи CoVG, гетерологичный белок, экспрессирующих ооцитов мембрана состоит из трех областей. Ионный ток данные записываются с верхней области ооцита какванна, окружающий верхнюю область крепится к командной потенциалом, который может быть легко и быстро изменить. Средней области защищает от токов утечки, будучи прикрепленными к тем же потенциалом, верхней области 9. В нижней области, где отверстие ооцитов (вырезать открыт) происходит с использованием раствора сапонина или канюли. Химические или ручное открывание мембраны в донной области позволяет управлять внутреннего потенциала, который крепится к земле, и оказывает внутрь клетки прилегает к нижней камере раствора. Перфузии решений в нижнюю камеру можно настроить свойства внутренней среды, в то время как обмен решение в верхней камере изменяет внешний антураж. 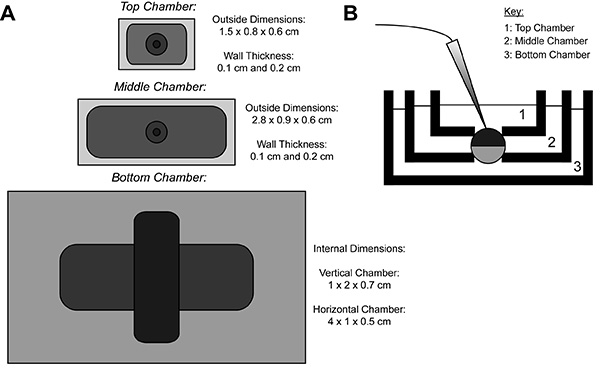
Рисунок 1. Ооцитов Cut-Открыть Напряжение-зажим Ванна Схема установки. () ТопВвиду вниз из трех ванн, разделенных друг от друга. Размеры камер для CoVG отображаются на рисунке. (B) Вид сбоку установки ванны в положении тестирования. Нажмите здесь, чтобы увеличить изображение .
Преимущества метода CoVG включают низкую текущую шум (1 нА при 3 кГц), контроль ионного состава внешней среды, в способности модулировать внутренний СМИ, разрешение быстро времени (20-100 мкс постоянная времени распада Емкость переходные), и стабильные записи в течение нескольких часов 9. К недостаткам можно отнести, что он требует специального оборудования и труднее выполнять по сравнению с двумя напряжения на электродах зажима (TEVC) 10.
В то время как подход CoVG требует высокой специализированное оборудование и сложные процедурные элементы, он может позволить для приобретения Valuспособные электрофизиологические данные. Эти данные, такие как стробирования токов с быстрой кинетики и хвостовые токи 4, могут быть записаны без некоторых проблем, связанных с другими протоколами напряжение зажима включая канала кратком изложении. Незначительные изменения в настройки CoVG может позволить для использования регуляторов температуры и зажим напряжение флуорометрии (VCF). Включение напряжения зажим элементов флуорометрии в сборке CoVG может увеличить выход данных присвоением способность контролировать белка конформационные изменения, одновременно записывая текущее 11-13.
