Técnicas de fijación de voltaje especializados permiten la grabación de corrientes iónicas en los potenciales de membrana controlados. Ampliamente utilizado de fijación de voltaje de dos electrodos (TEVC) y las técnicas de patch clamp proporcionan información electrofisiológica fiable sobre las propiedades de muchos canales iónicos. Sin embargo, ambos de estos métodos tienen inconvenientes que impiden la adquisición de datos fiables para los canales de sodio dependientes de voltaje rápidos y otros canales de activación rápida en membranas tales como las de los oocitos de Xenopus. Los laboratorios Bezanilla y Stefani consecuencia desarrollaron la técnica de fijación de voltaje brecha vaselina a cielo abierto (COVG) de ovocitos 2. La técnica se ha aplicado ampliamente para grabar, Na +, K + y Ca 2 + canales 3-8.
Durante la grabación COVG, una membrana del oocito que expresa la proteína heteróloga se divide en tres regiones. Los datos de corriente iónica se graba desde la región superior de la ovocito como labaño que rodea la región de la parte superior se sujeta a un potencial de comando, que se puede cambiar fácil y rápidamente. Los guardias región de Oriente contra corrientes de fuga al ser sujetado al mismo potencial que la región superior 9. La región inferior es que la apertura de los ovocitos (cut-open) se produce mediante el uso de una solución de saponina o una cánula. Químico o la apertura manual de la membrana en la zona del fondo permite el control del potencial interno, que está fijado a tierra, y hace que el interior de la célula contigua con la solución de cámara inferior. La perfusión de soluciones en la cámara inferior puede ajustar las propiedades del medio interno, mientras que el intercambio de solución en la cámara superior altera el entorno externo. 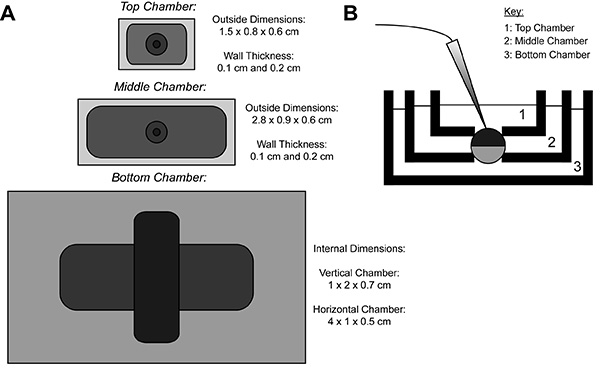
Figura 1. Ovocitos-Cut abierto Voltaje-Clamp Bath esquema de instalación. (A) Topabajo de la vista de los tres cuartos de baño separados el uno del otro. Las dimensiones de las cámaras para COVG se muestran en la figura. (B) Vista lateral de la configuración de los baños en posición de prueba. Haga clic aquí para ver la imagen más grande .
Las ventajas de la técnica de COVG incluyen bajo ruido de corriente (nA 1 a 3 kHz), el control de la composición iónica de los medios de comunicación externos, la capacidad de modular los medios de comunicación interna, el tiempo de resolución rápida (20-100 microsegundos constante de tiempo de decaimiento de la capacidad transitoria), y grabaciones estable durante varias horas 9. Las desventajas son que requiere un equipo especializado y es más difícil de realizar en comparación con dos de fijación de tensión de electrodos (TEVC) 10.
Si bien el enfoque COVG requiere un equipo altamente especializado y elementos de procedimiento intrincados, puede permitir la adquisición de valiosadatos electrofisiológicos capaces. Estos datos, como conmutar corrientes con cinética rápida y corrientes de cola 4, se puede grabar sin algunos de los problemas asociados con otros protocolos de fijación de voltaje, incluyendo canal de decadencia. Pequeñas modificaciones en la configuración COVG pueden permitir el uso de controladores de temperatura y fluorometría fijación de voltaje (VCF). La inclusión de elementos de sujeción de fluorometría de tensión dentro del conjunto COVG puede aumentar la salida de datos por conferir la capacidad de monitorizar los cambios conformacionales de la proteína mientras que al mismo tiempo la grabación actual 11-13.
