As células estaminais adultas (SCS) são essenciais para a manutenção da homeostase do tecido, substituindo células que morrem e reparar tecidos danificados após a lesão. Estes SCs são definidas pela sua capacidade para sofrer auto-renovação contínua e de se diferenciar em linhagens celulares diferentes 1-3. Os sistemas mais bem estudados, as quais são dependentes de SCs adultos para a sua reposição, incluem o sistema hematopoiético, o intestino e o 1,2,4 pele.
Durante a embriogénese, a pele começa como uma única camada de células epidérmicas. Morfogênese do folículo piloso (HF) começa quando as células mesenquimais preencher a pele e formam uma subjacentes derme colágenas 5. Especializada células mesenquimais, que mais tarde constituem a papila dérmica (DP), organizam diretamente abaixo da camada epidérmica e estimulam o epitélio para formar placodes cabelo que começam a crescer para baixo 6. Altamente proliferação de células da matriz, situados na parte inferior do HF,envelope estas células mesenquimais e formar o bulbo capilar, enquanto que a camada interna começa a diferenciar-se em cilindros concêntricos de modo a formar o eixo do cabelo (HS) e a bainha envolvente interior raiz (IRS) 2,3.
Na vida pós-natal a epiderme da pele é composta de três compartimentos: a epiderme interfoliculares (IFE), a glândula sebácea (SG) e o HF. Em contraste com o IFE e SG, que estão em constante estado de homeostase, o HF é um mini-órgão dinâmico que sofre ciclos contínuos de crescimento (anágena), destruição (catágena) e repouso (telógena) 4,7. As células-tronco do folículo piloso (HFSCs) que o combustível este ciclo perpétuo, residir em um nicho especializado dentro do HF conhecido como o bojo 4. Durante anágena os HFSCs sair do bojo, seguindo sinais de ativação do DP, começam a proliferar e descer para baixo, criando assim uma trilha linear longo de células conhecidas como a bainha externa da raiz (ORS) 8-10. As células da matriz, quecercar o DP na base de HF, rapidamente ciclo e migrar para cima passando por diferenciação terminal gerando assim o HS e o IRS 10 (Figura 1). A duração do anágeno determina o comprimento do cabelo e é dependente da capacidade proliferativa e de diferenciação das células da matriz 6. Quando o HF entra catágena, as células da matriz-amplificação de trânsito na cessar lâmpada a proliferar, sofrem apoptose e regridem completamente enquanto puxa o DP para cima até que ele atinja a parte não-ciclismo do HF 8,11. Durante esta retracção, o HF forma uma estrutura temporário conhecida como a cadeia epitelial, que é característica de catágena e contém muitas células apoptóticas. Em ratinhos, catágena dura entre 3-4 dias e é altamente sincronizadas no primeiro ciclo do cabelo. Quando o HF atinge todos os telógena HFSCs se tornar quiescente. As diferentes etapas do ciclo de IC também são caracterizadas por alterações na cor da pele do rato devido a mprodução elanin. As alterações na pele de preto durante anágena ao cinza escuro durante catágena a rosa durante telógeno 6,7,12,13.
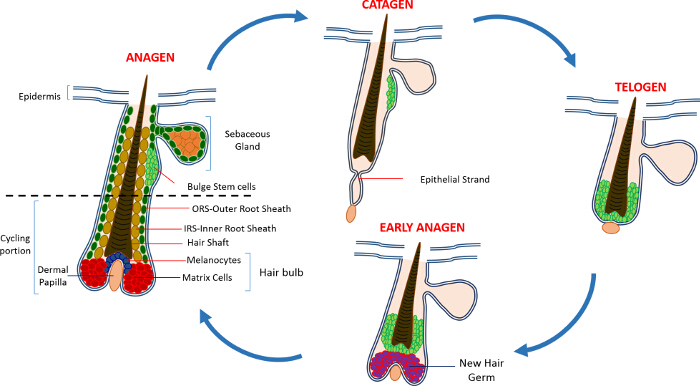
Figura 1: O ciclo de folículo piloso. O HF é composto por uma parte superior permanente e um menor em constante remodelação, a porção de ciclismo que sofre ciclos contínuos de crescimento rápido (anágena), destruição (catágena) e uma fase de quiescência parente ou repouso (telógena). Por favor clique aqui para ver uma maior versão desta figura.
Os SCs mantendo a HF foram inicialmente identificados utilizando experimentos de perseguição, com timidina tritiada, que revelaram uma população de células de retenção lenta etiqueta ciclismo (LRC) que residiam na região permanente do HF logo abaixo da SG 14. Avanços na HFSCCaracterização revelou um pequeno número de marcadores que podem ser utilizados para identificar e isolar SCs específicos do nicho da IC 15. Talvez o melhor marcador para o enriquecimento de células CD34 é HFSCs, um marcador de superfície celular também identificado como um marcador SC hematopoiéticas em seres humanos 16. Dentro deste CD34 + populações duas populações distintas também foram isoladas com base na expressão da integrina α6 2. Outro marcador é queratina 15 (K15), que é altamente expresso na região da protuberância, co-localiza com a expressão de CD34 e um promotor K15 é usado para o direccionamento e isolando HFSCs em animais transgénicos 15,17-19. Na década passada várias outras populações distintas de células progenitoras HFSCs e também têm sido relatados para residir dentro do IC 17,20-27.
Um recurso interessante adicional de HFSCs é a sua contribuição para a reparação da pele. Em condições normais HFSCs reabastecer o HF e não tomar parte em IFE homeostase. HoweVer, em resposta ao ferimento, estas células sair do seu nicho SC e ajuda na repovoamento a IFE 9. Nós demonstramos recentemente que os ratinhos excluído para a exibição do gene SEpt4 / ARTE pró-apoptótica um aumento do número de células CD34, K15 e Sox9 + HFSCs, que demonstram uma resistência à apoptose. HFSCs foram isolados a partir SEpt4 / ARTE – / – peles dorsal utilizando células activadas por fluorescência (FACS) e não havia mais do que um aumento de duas vezes no número de células CD34 + e + K15 HFSCs. Estes SEpt4 / ARTS – / – HFSCs foram expandidas in vitro e não só deu origem a mais colónias, mas também foram capazes de suportar as condições mais severas, em comparação com controles 28.
Como resultado de ter um maior número de HFSCs, SEpt4 ARTE / – / – ratos curados significativamente mais rapidamente em resposta a lesões excisão de pele. Surpreendentemente, SEpt4 / Artes – / – ratos displayeda grande número de HF regenerado a partir do leito da ferida, cicatrizes e significativamente menores. Além disso, os ratos excluídos para XIAP (inibidor ligada ao X de apoptose), o alvo bioquímico de ARTS, demonstrou prejudicou a cicatrização 28.
Os nossos resultados e trabalho realizado em outros laboratórios mostraram que HFSCs servir como um modelo ideal para o estudo da biologia e função de SCs adultos. Aqui, nós descrevemos a metodologia para o enriquecimento e isolamento de HFSCs e queratinócitos epidérmicos com base na expressão de quatro marcadores: α6 integrina; β1 integrina; Sca-1 (um marcador para queratinócitos epidérmicos) e CD34. Semelhante isolamento de K15 + HFSCs também pode ser realizada utilizando o rato K15 repórter GFP-19.
