Взрослые стволовые клетки (SCS) необходимы для поддержания гомеостаза тканей путем замены умирающие клетки и восстановления поврежденных тканей после повреждения. Эти Стволовые определяются их способности претерпевать постоянному самообновлению и дифференцироваться в различные клеточные клоны 1-3. Наиболее изучены системы, которые зависят от взрослых ГКС для их пополнения, включают в себя систему гемопоэза, кишечник и 1,2,4 кожи.
Во время эмбриогенеза, кожа начинает как один слой эпидермальных клеток. Морфогенез волосяного фолликула (HF) начинается , когда мезенхимальные клетки заселить кожу и образуют основные коллагеновые дермы 5. Специализированные мезенхимальные клетки, которые впоследствии составляют сосочков дермы (DP), упорядочивать непосредственно под эпидермальным слоем и стимулируют эпителий плакоды , чтобы сформировать волосы , которые начинают расти вниз 6. Высоко пролиферирующие матричные клетки, расположенные в нижней части ВЧ,конверт эти мезенхимные клетки и образуют луковицы, в то время как внутренний слой начинает дифференцироваться в концентрических цилиндров , чтобы сформировать волос вала (HS) и окружающий внутреннюю оболочки корня (IRS) 2,3.
В постнатальной жизни эпидермис кожи состоит из трех отделения: интерфолликулярной эпидермис (IFE), сальные железы (SG) и HF. В отличие от IFE и SG , которые находятся в постоянном состоянии гомеостаза, ВЧ представляет собой динамичный мини-орган , который претерпевает непрерывные циклы роста (анагена), уничтожение (катагена) и покоя (телоген) 4,7. Волосяной фолликул стволовые клетки (HFSCs) , что топливо , этот вечный цикл, постоянно находиться в специализированной нише в пределах HF , известной как выпуклость 4. Во время анагена в HFSCs выхода из выпуклость, следующие сигналы активации с DP, начинают пролиферировать и спускаемся вниз , таким образом , создавая длинный линейный след клеток , известных как внешней оболочки корня (ПРС) 8-10. Матричные клетки, которыеокружают DP на базе ВЧ, быстро цикла и мигрировать вверх проходит терминальной дифференцировки , таким образом , генерирующее HS и IRS 10 (рисунок 1). Продолжительность анагена определяет длину волос и зависит от пролиферативного и дифференциации мощности матричных ячеек 6. Когда HF входит катагена, транзитно-усилительных матричные клетки в колбы перестают размножаться, подвергаются апоптозу и регрессируют полностью, потянув DP вверх , пока он не достигнет , не задействуя часть ВЧ 8,11. Во время этого втягивания ВЧ образует временную структуру, известную как эпителиальный нить, которая характерна для катагена, и содержит много апоптозных клеток. У мышей, катаген длится от 3-4 дней и высокосинхронной в первом цикле волос. Когда HF достигает телоген все HFSCs становятся в состоянии покоя. Отличительными этапы цикла HF также характеризуются изменением цвета кожи мыши вследствие мelanin производство. Изменения кожи от черного во время анагена до темно – серого цвета во время катагена до розового во время телогене 6,7,12,13.
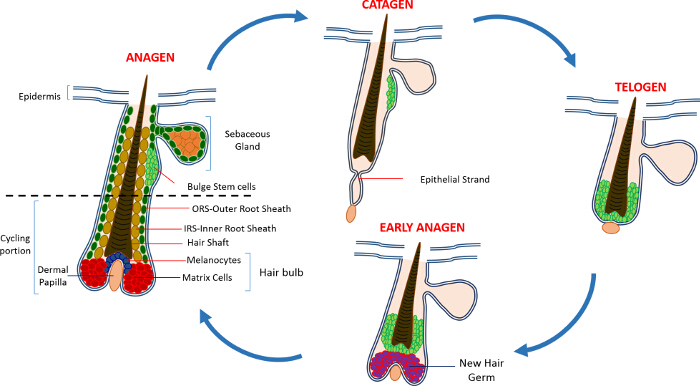
Рисунок 1: волосяной фолликул цикл. КВ состоит из постоянной верхней части и нижней постоянно ремоделирования, езда на велосипеде часть , которая претерпевает непрерывные циклы быстрого роста (анагена), уничтожение (катагена) и относительной фазы форсированном или покоя (телоген). Пожалуйста , нажмите здесь , чтобы увидеть увеличенную версия этой фигуры.
ГКС поддержание ВЧ первоначально были идентифицированы с использованием чейза экспериментов, с помощью меченного тритием тимидина, которые показали население медленно меток задействуя удерживающих клеток (LRC) , которые проживали в постоянной области HF чуть ниже SG 14. Достижения в HFSCхарактеристика выявлено небольшое количество маркеров , которые могут быть использованы для идентификации и выделения конкретных SCs из HF ниши 15. Возможно, лучший маркер для обогащения HFSCs является CD34, поверхностный маркер клеток также идентифицирован как кроветворной SC маркера в организме человека 16. В пределах этой популяции CD34 + две различные группы населения также были выделены основаны на α6 экспрессии интегринов 2. Другой маркер является кератин 15 (К15) , который экспрессируется на высоком уровне в области выпуклость, совместно локализуется с экспрессией CD34 и К15 промотор используется для ориентации и выделения HFSCs в трансгенных животных 15,17-19. В последнее десятилетие несколько других различных популяций HFSCs и клеток – предшественников также сообщалось проживать в пределах HF 17,20-27.
Еще интересной особенностью HFSCs является их вклад в ремонт кожи. При нормальных условиях HFSCs пополнения HF и не принимают участия в IFE гомеостаза. Хаувер, в ответ на язву эти клетки выйдут из SC ниши и помощь в заселив ФИИ 9. Недавно мы показали , что у мышей удалены для проапоптотического дисплея гена Sept4 / ARTS повышенное количество CD34, K15 и Sox9 + HFSCs, которые демонстрируют устойчивость к апоптозу. HFSCs были выделены из Sept4 / ИСКУССТВ – / – спинные шкуры с использованием флуоресценции активированной сортировки клеток (FACS) и там было больше , чем в два раза больше числа CD34 + и K15 + HFSCs. Эти Sept4 / ARTS – / – HFSCs были расширены в пробирке и не только привели к более колоний , но также способны выдерживать более жесткие условия по сравнению с контрольной группой 28.
В результате того , что повышенное количество HFSCs, Sept4 / ARTS – / – мышей значительно быстрее зажила в ответ на иссечение кожи травм. Поразительно, Sept4 / ARTS – / – мышей displayeда большое количество регенерированных HFs из раневого ложа, а также значительно меньшие шрамы. Кроме того, мыши , удаленные для XIAP (ингибитор Х-хромосомой апоптоза), биохимической мишени ИСКУССТВ, продемонстрировали ухудшение заживления 28.
Наши результаты и работы, выполненные в других лабораториях, показали, что HFSCs служат идеальной моделью для изучения биологии и функции взрослых SCS. Здесь мы описываем методологию для обогащения и выделения HFSCs и эпидермальных кератиноцитов на основе экспрессии четырех маркеров: интегрин а6; интегрин β1; Sca-1 (маркер для кератиноцитов эпидермиса) и CD34. Подобное выделение K15 + HFSCs также может быть выполнена с использованием K15-GFP – репортер мыши 19.
