وهناك سمة مشتركة من اضطرابات وأمراض الجهاز العصبي المركزي، مثل إصابات الدماغ الصدمة (تبي)، إصابة الحبل الشوكي (سسي) والسكتة الدماغية ومرض الزهايمر، ومرض باركنسون، هو انقطاع مسارات محور عصبي والخلايا العصبية الخسارة 1 ، 2 ، 3 ، 4 ، 5 ، 6 . على سبيل المثال، عندما يذهب السكتة الدماغية دون علاج، ويقدر أن محاور يتم فقدان بمعدل 7 أميال من المحاور في الدقيقة 5 . في حالة تبي، والتي ما يقرب من 1.7 مليون شخص تجربة كل عام في الولايات المتحدة وحدها، قد تستمر انحطاط محور عصبي تحدث بعد سنوات من الصدمة، كما يعجل الإصابة الأولية على المدى الطويل الدولة الاعصاب 4 . ومما يزيد من حدة هذه الآثار الضارة أن الجهاز العصبي المركزي لديه كابا محدود للغايةمدينة للتجديد 1 ، 7 ، 8 ، 9 . بعد الإصابة، وتطور البيئة المثبطة التي تتميز بعدم توجيه توجيهات إلى أهداف بعيدة، وجود مثبطات المايلين المرتبطة التي تعيق نمو العصبية، وتشكيل ندبة الدبقية من قبل الخلايا النجمية رد الفعل 8 ، 10 ، 11 ، 12 . الندبة الدبقية بمثابة الحاجز البيوكيميائية والفيزيائية لتجديد، مع جزيئات مثل بروتيوغليكان كبريتات شوندروتن عرقلة نمو محور عصبي 8 ، 11 . وعلاوة على ذلك، على الرغم من أن الخلايا الجذعية العصبية تم العثور عليها في الجهاز العصبي المركزي الكبار، وإنتاج الخلايا العصبية الجديدة محدودة، ودليل ثابت على تكوين الخلايا العصبية لم يتم العثور عليها إلا في لمبة الشم، والحصينالمنطقة تحت الحبيبية، ومنطقة المحيطة بالبطين، والقناة المركزية للحبل الشوكي 13 ، 14 . هذه العقبات تمنع الانتعاش وظيفية من الخلايا العصبية المفقودة والهندسة المعمارية المادة البيضاء بعد الإصابة أو المرض، مما أدى إلى تغير الحياة في كثير من الأحيان وطويلة الأمد لهذه الظروف.
على الرغم من عدم وجود القدرة التجدد في الجهاز العصبي المركزي الكبار، وقد ثبت أن تجديد محور عصبي ممكن إذا تم تقديم الإشارات البيئية الكافية لاستضافة الخلايا العصبية 15 ، 16 ، 17 ، 18 . وقد حاول الباحثون تقديم ومعالجة عوامل النمو (على سبيل المثال، عامل نمو الأعصاب ، عامل نمو البشرة، عامل النمو المعتمد على الدبقية، والعامل العصبي -3) وغيرها من جزيئات التوجيه لتحفيز اللدونة وتجديد المحوار 14 ،/ سوب> 18 ، 19 . على الرغم من أن هذه الدراسات أكدت أن محاور الكبار قادرة على الاستجابة لعوامل النمو، وهذه الاستراتيجيات محدودة بسبب نفاذية منخفضة من حاجز الدم في الدماغ والتدرجات المكانية والزمانية المحددة المطلوبة لتعزيز التجدد 14 ، 18 ، 19 . وقد اعتمدت نهج أخرى على فرط تنشيط عوامل النسخ ذات الصلة التجدد في الخلايا العصبية الجهاز العصبي المركزي. على سبيل المثال، أوفيركسريسيون من عامل النسخ ST3 حفز تجديد محور عصبي في العصب البصري 20 . ومع ذلك، كلا تسليم جزيء حيوي و أوفيركسريسيون من عوامل النسخ تفشل في استبدال السكان العصبية المفقودة. وقد تركزت الاستراتيجيات القائمة على الخلية أساسا على زرع الخلايا الجذعية العصبية (نسس) في الجهاز العصبي المركزي، والاستفادة من قدرتها على استبدال الخلايا العصبية الجهاز العصبي المركزي، والإفراج عن العوامل الغذائية،ودعم محاولات العصبية التي تحدث بعد الإصابة 17 . على الرغم من هذا، لا تزال هناك تحديات ملحة تعوق هذا النهج، بما في ذلك القدرة المعوقة للخلايا العصبية المزروعة من أجل البقاء، والاندماج مع المضيف، وتظل مقيدة مكانيا إلى المنطقة المصابة 6 ، 14 ، 17 ، 21 . وبالإضافة إلى ذلك، تسليم الخلية وحدها غير قادرة على إعادة الهندسة المعمارية الخلوية من مسارات محور عصبي التالفة أو المفقودة. وهناك نهج بديل يعالج المشاكل التي تواجه استراتيجيات تسليم الخلايا والمخدرات / الكيميائية هو الجمع بين هذه النهج مع استخدام المواد الحيوية 14 ، 22 ، 23 . المواد الحيوية مثل الهلاميات المائية هي قادرة على محاكاة الخصائص البيوكيميائية والفيزيائية للمصفوفة خارج الخلية (إسم)، والمساعدة في تسليم الخليةد داخل المنطقة المصابة، وتسليم عوامل النمو وغيرها من الجزيئات النشطة بيولوجيا مع الافراج عن السيطرة 22 . وقد أدت الخصائص الجذابة لهذه الاستراتيجيات القائمة على المواد الحيوية في أدلة على الجسم الحي تجديد محور عصبي بعد زرع السقالات إلى منطقة ليسيونيد 24 ، 25 ، 26 ، 27 ، 28 ، 29 ، 30 . ومع ذلك، واستراتيجيات المواد الحيوية الخلوية لا تحل محل السكان العصبية المفقودة. عندما تستخدم كوسيلة تسليم الخلايا العصبية، الدبقية، أو الخلايا العصبية السلائف، والمواد الحيوية غير قادرة على إعادة تشكيل شبكات محور عصبي لمسافات طويلة. التحدي المتمثل في تطوير النهج الذي يعالج كل من محور عصبي انحطاط المسار وفقدان الخلايا العصبية المرتبطة إصابة الجهاز العصبي المركزي والمرض لا يزال <سوب كلاس = "كريف"> 31.
وقد ذكرت مجموعة أبحاثنا في وقت سابق تطوير الشبكات العصبية زرع الأنسجة العصبية زرع (تينس الصغرى)، والتي هي نوع من "سقالة المعيشة" تتكون من الهيئات خلية الخلايا العصبية تقتصر على واحد أو طرفي أغاروس هيدروجيل-إسم الصغرى العمود ، مع محاذاة محاور عصبية تمتد في جميع أنحاء المناطق الداخلية من هذا ثلاثي الأبعاد (3D) التشفير 1 ، 10 ، 31 ، 32 . واحدة من الاختلافات الرئيسية بين هذه التقنية والنهج السابقة هو أن الهندسة المعمارية الخلوية من تينس الصغرى يتم إنشاؤها تماما في المختبر وزرع بعد ذلك 33 ، 34 ، 35 ، 36 ، 37 ، 38 ، <سوب كلاس = "كريف"> 39 ، 40 ، 41 . في التصنيع المختبر يوفر السيطرة المكانية والزمانية واسعة من النمط الظاهري الخلوي والتوجه، والخصائص الميكانيكية / الفيزيائية، والإشارات البيوكيميائية، والعوامل الخارجية، والذي يفيد دمج هذه السقالات مع المضيف بعد زرع 41 ، 42 . الصغرى تينس مستوحاة تشريحيا لأنها تحاكي الدماغ العصبي، وعرض المسالك محور عصبي مماثلة لتلك التي تجسر مناطق وظيفية متميزة من الدماغ ( الشكل 1A ) 1 . لذلك، قد تكون هذه الاستراتيجية قادرة على استبدال جسديا مساحات المادة البيضاء المفقودة والخلايا العصبية بعد زرع في منطقة ليسيونيد. هذا الأسلوب هو أيضا مستوحاة من الآليات التنموية التي "السقالات الحية الطبيعية" التي شكلتها الخلايا الدبقية شعاعي والمحاور عصبية رائدة بمثابة أدلة دالة للخليةوالهجرة من المنطقة تحت البطينية ونمو محور عصبي، على التوالي 43 . يتم تلخيص هذه الآليات في محاور محور عصبي الانحياز من تينس الصغرى، والتي يمكن أن تقدم مسارات حية للهجرة الخلايا العصبية وتجديد محور عصبي من قبل محور عصبي بوساطة محور عصبي ( الشكل 1C ) 43 . وعلاوة على ذلك، هذه الاستراتيجية تستفيد من التكامل متشابك بين الخلايا العصبية تين الصغرى والدوائر الأصلية، وتشكيل التبديلات الجديدة التي قد تسهم في الانتعاش وظيفية ( الشكل 1B ) 43 . القدرة على تشكيل المشبك قد تمنح أيضا هذا النهج القدرة على تعديل الجهاز العصبي المركزي والرد على الأنسجة المضيف وفقا لتغذية مرتدة الشبكة. على سبيل المثال، يمكن تحفيز الخلايا العصبية النشطة أوبتوجينيتيكالي في السقالات الحية لتعديل الخلايا العصبية المضيف من خلال التفاعلات متشابك ( الشكل 1D ).
وبالإضافة إلى ذلك، فإن كونستر أنبوبي القائم على المواد البيولوجيةأوكتيون من تينس الصغرى يوفر بيئة كافية لالتصاق الخلايا، والنمو، وتمديد نيوريت، والإشارات، في حين أن أبعاد مصغرة من يبني يمكن أن تسمح غرس زرع الحد الأدنى، وتوفير المكروية جزئيا عزلها للاندماج التدريجي في الدماغ. في الواقع، وقد أظهرت المنشورات الأخيرة إمكانات تينس الصغرى لمحاكاة المسارات العصبية بعد زرع في الدماغ الفئران. وبعد حقن مكروي التجسيمي، ذكرنا سابقا أدلة على البقاء على قيد الحياة الصغرى تين الخلايا العصبية، وصيانة الهندسة المعمارية محور عصبي، وتوسيع العصبية في القشرة المضيف إلى 1 شهر على الأقل في الجسم الحي 10 ، 31 . وعلاوة على ذلك، وضع العلامات مع سينابسين قدمت أدلة نسيجية من التكامل متشابك مع الأنسجة الأصلية 10 ، 31 . وعموما، يمكن أن تكون شبكات تينس الصغرى مناسبة بشكل فريد لإعادة بناء وتعديل التالفنس عن طريق استبدال الخلايا العصبية المفقودة، ودمج متشابك مع الدوائر المضيف، واستعادة المفقودة الهندسة المعمارية المحاور المفقودة، وفي بعض الحالات، وتوفير محاور عصبية تجديد مع العظة المسارات المناسبة.
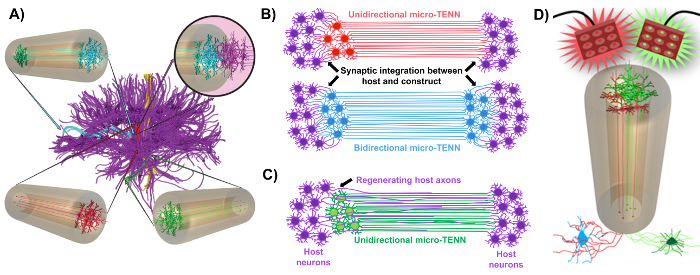
الشكل 1: المبادئ والإلهام وراء تطوير شبكات العصبية الأنسجة الدقيقة المهندسة (تينس الصغرى). ( A ) الصغرى تينس تقليد الهندسة المعمارية الخلوية من الدماغ كونكتوم (الأرجواني)، التي ترتبط مناطق متميزة وظيفيا من قبل طويلة محاذاة محاور عصبية في اتجاه أحادي الاتجاه (الأحمر والأخضر) أو ثنائية الاتجاه (الأزرق). على سبيل المثال، يمكن لشبكات تين الصغرى إعادة تكوين الوصلات المفقودة في المسالك القشرية والنيغروستريتية أو في المسار الثاقب من القشرة المخية إلى الحصين (مقتبس من ستروزينا وآخرون ، 2015) 1 . ( B ) رسم تخطيطي ل أونيديركتيونال و ثنائية الاتجاه الصغرى تين (الأحمر والأزرق، على التوالي) دمج متشابك مع الدوائر المضيف (الأرجواني) لتكون بمثابة تتابع وظيفي بين طرفي الآفة. ( C ) تخطيطي من المسالك المحورية من أحادي الاتجاه تين الصغرى (الأخضر) بمثابة دليل لتجديد يسرع محوار عصبي محاور (الأرجواني) نحو الهدف الذي تتفاعل الصغرى تين. ( D ) الرسم التخطيطي المفاهيمي لاستخدام تينس-أوبتوجينيتيكالي الصغرى تينس كما نيورومودولاتورس، والاستفادة من التكامل متشابك مع الخلايا العصبية استثارة أو مثبطة (القاع). الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
المخطوطة الحالية تفاصيل المنهجية المستخدمة في تلفيق الصغرى تينس باستخدام الخلايا العصبية القشرية الدماغية المستمدة من الجنين. ومن الجدير بالذكر أن تينس الصغرى يمكن أن تكون ملفقة مع أنواع أخرى من الخلايا العصبية. على سبيل المثالوافرة، وتقارير أولية ناجحة التنمية الدقيقة الصغرى ظهرت الظهري العقدة الجذرية العقدة (درغ) الخلايا العصبية 32 . يمكن توليد الأعمدة الصغيرة هيدروجيل ( الشكل 2A ) عن طريق إضافة الاغاروز السائل إلى صفيف قناة أسطواني مصنوع حسب الطلب، أو أنابيب الشعرية، على حد سواء تحتوي على محاذاة إبر الوخز بالإبر. الإبرة تشكل التجويف ويحدد القطر الداخلي (إد) من العمود الصغير، في حين أن معرف أنبوب الشعرية وقطر الاسطوانات في جهاز قطع الليزر تملي القطر الخارجي (أود) من يبني. يمكن اختيار أود و إد وفقا للتطبيق المطلوب عن طريق اختيار أقطار مختلفة للجهاز / أنابيب الشعرية وإبر الوخز بالإبر، على التوالي. ويمكن أيضا أن يتغير طول الأعمدة الصغيرة. حتى الآن، أبلغنا عن بناء الصغرى تينس تصل إلى 20 ملم في الطول 10 وتسعى بنشاط حتى أطوال أطول. بعد المواد الهلامية الاغاروز والوخز بالإبر نتتم إزالة إيدلز، يتم إضافة حل إسم تتألف عموما من نوع I الكولاجين و لامينين إلى التجويف من يبني ( الشكل 2C ). يوفر جوهر إسم سقالة لدعم التصاق الخلايا العصبية ونمو محور عصبي. في البداية، كانت الخلايا العصبية القشرية الفئران الأولية مطلي في الأعمدة الصغيرة باستخدام تعليق خلية فصل 10 ، 31 ، 32 . ومع ذلك، فإن هذا النهج لم تنتج الهندسة المعمارية المستهدفة في جميع الحالات، والتي تم تعريفها على أنها الهيئات الخلية العصبية يقتصر على نهايات الأعمدة الصغيرة، مع التجويف المركزي تتألف من محض محاذاة محاور العصبية. ومنذ ذلك الحين، وقد مكن استخدام طريقة التجميع العصبي القسري (على أساس البروتوكولات تكييفها من أونغرين وآخرون ) تصنيع أكثر موثوقية ومتسقة من تينس الصغرى مع هيكل مثالي ( الشكل 2B ) 44 . بالإضافة إلى وصف التيارالمنهجية، هذه المادة سوف تظهر ممثل على النقيض من المرحلة والصور متحد البؤر من تينس الصغرى التي تثبت تشكيل المسالك محور عصبي مع مرور الوقت، فضلا عن الهدف النهائي الهندسة المعمارية. وهذه المخطوطة توسيع أيضا على الجوانب الجديرة بالملاحظة من البروتوكول والتحديات المتبقية والاتجاهات المستقبلية للتكنولوجيا تين الصغرى.
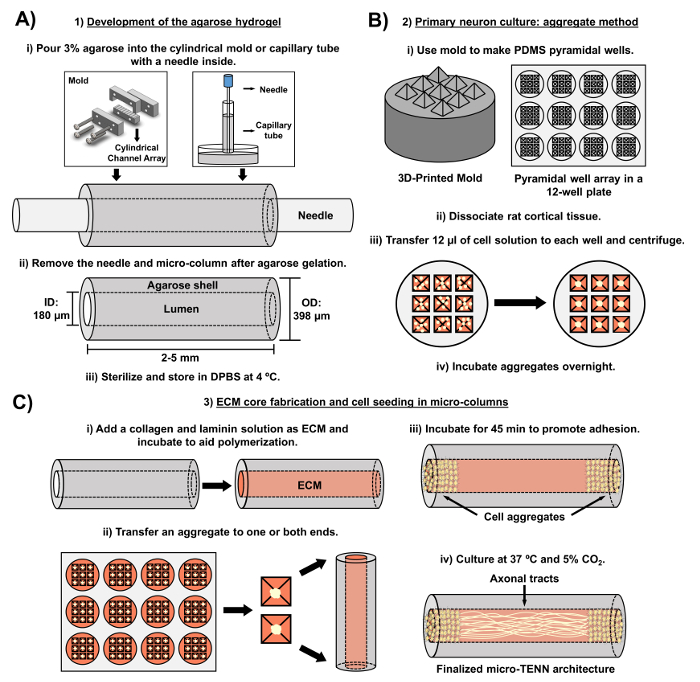
الشكل 2: الرسم التخطيطي لعملية التصنيع الصغرى تين ثلاث مراحل. ( أ ) تطوير هيدروجيل الاغاروز: (1) في البداية، يتم إدخال إبرة الوخز بالإبر الصغيرة (على سبيل المثال ، 180-350 ميكرون في القطر) في القنوات اسطوانية من صنع خصيصا، وقص الليزر قطع أو أنبوب شعري (على سبيل المثال ، 380-700 ميكرون في القطر). في الخطوة التالية، يتم عرض الاغاروز السائل في دبس في القنوات اسطوانية أو أنابيب الشعرية. (إي) بعد المواد الهلامية الاغاروز، تتم إزالة الإبرة ويتم تفكيك القالب لانتاج أجاروس الأعمدة الصغيرة جوفاء. (3) يتم بعد ذلك تعقيم هذه البنايات وتخزينها في دبس. ( ب ) ثقافة الخلايا العصبية الأولية وطريقة التجميع: (1) يتم تنفيذ تجميع الخلايا العصبية في صفيف هرمي الصغرى جيدا، يلقي من قوالب 3D المطبوعة، التي تناسب في آبار لوحة الثقافة 12 جيدا. (2) تشمل الشبكات الصغرى تينس الخلايا العصبية الفئران الأولية فصلها عن أدمغة الجنين من الجنينية-يوم 18 الجرذان. بعد تفكك الأنسجة مع التربسين إدتا ودناز الأول، يتم إعداد محلول خلية مع كثافة 1.0-2.0 × 10 6 خلية / مل. (إي) يتم نقل 12 ميكرولتر من هذا الحل إلى كل بئر في مجموعة هرم الصغيرة أيضا. يتم طرد لوحة تحتوي على هذه الآبار الصغيرة لإنتاج المجاميع الخلية. (4) ثم يتم احتضان هذه بين عشية وضحاها قبل الطلاء في الأعمدة الصغيرة. ( C ) إسم تلفيق الأساسية والبذر الخلية: (ط) قبل زرع الخلايا، والحل إسم تحتوي على 1 ملغ / مل نوع الأول الكولاجين و 1 ملغ / مليتم نقل لامينين إلى المناطق الداخلية من تينس الصغرى ويسمح للبلمرة. (2) اعتمادا على ما إذا كان يتم تصنيع تينس أحادي الاتجاه أو ثنائي الاتجاه تينس، يتم وضع مجمع في واحد أو كليهما من العمود الصغير، على التوالي. (3) بعد فترة من الحضانة لتعزيز الالتصاق، يتم زراعة تينس الصغرى في أطباق بتري غمرت مع المستمدة الجنينية المتوسطة القاعدية العصبية. (4) بعد 3-5 أيام في الثقافة، وينبغي أن الهيكل النهائي تين الدقيقة تظهر مجاميع الخلايا في أقصى النقيض من العمود الصغير، مع مساحات محور عصبي تمتد طوله. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
