Nonostante i vantaggi offerti da biocatalisi, l’integrazione di biocatalitici passi nelle vie sintetiche o rotte biocatalitici totale rimane per lo più limitata a risoluzioni di cinetiche enzimatiche. Questi percorsi sono stati ampiamente utilizzati come un primo passo nella sintesi asimmetrica chemo-enzimatica, ma biocatalisi offre molte più possibilità nel gruppo funzionale interconversioni con elevata stereoselettività1,2,3 . Inoltre, come reazioni biocatalitici sono condotte in condizioni simili, pertanto è possibile eseguire reazioni a cascata della moda uno-pentola4,5.
Amino alcoli chirali sono molecole versatili per l’utilizzo come ausiliari o impalcature in sintesi organica6. La frazione di alcool amminico è trovata frequentemente in metaboliti secondari e ingredienti farmaceutici attivi (API). Alcoli primari β-amminici sono prontamente disponibili dagli acidi α-amminici corrispondenti di sintesi chimica convenzionale, ma l’accesso a γ-amino alcoli chirali o amminoalcoli secondari richiede spesso noiose vie sintetiche insieme sensibile controllo della stereochimica7,8,9,10. A causa della sua elevata stereoselettività, biocatalisi possono fornire un percorso sintetico superiore a questi blocchi chirali11,12,13,14.
Precedentemente abbiamo segnalato la sintesi di mono – e di-idrossi-L-lisine di diastereoselettiva idrossilazione enzimatica catalizzata da diossigenasi del ferro (II) / α-chetoacido-dipendente famiglia ossigenasi (αKAO) (Figura 1)15. In particolare, a partire da L-lisina, la KDO1 dioxygenase catalizza la formazione di (3S) – derivato idrossilato (1), mentre il (4R) – derivato (2) è costituito dalla reazione con KDO2 diossigenasi. Regiodivergent successivi idrossilazioni da KDO1 e KDO2 portano alla formazione dei (3R, 4R) – diidrossi – L-lisina (3) in forma otticamente puro. Tuttavia, la gamma limitata di substrato di questi enzimi impedisce loro ampio utilizzo in sintesi chimica, soprattutto nell’idrossilazione di ammine semplici, come una molecola di acido carbossilico nella α-posizione del gruppo amminico è essenziale per attività16.
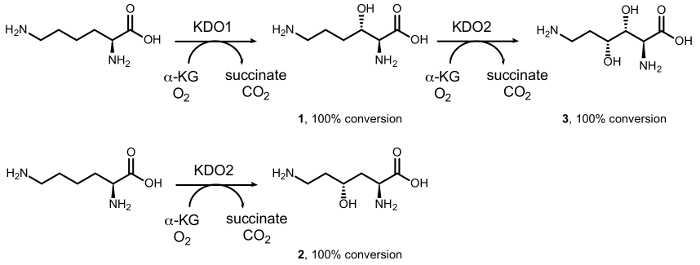
Figura 1: biocatalitici conversioni di L-lisina. Conversione in (3S) – idrossi – L-lisina (1) catalizzata da KDO1 dioxygenase; (4R) – idrossi – L-lisina (2) catalizzata da KDO2 dioxygenase; e (3R, 4R) – diidrossi – L-lisina (3) da reazione a cascata catalizzata successivamente da KDO1 e KDO2 diossigenasi. Clicca qui per visualizzare una versione più grande di questa figura.
Decarbossilazione è una reazione comune nel metabolismo17. In particolare, dell’amminoacido DCs (EC 4.1.1) sono privo di cofattore (piruvoil-dipendente) o enzimi PLP-dipendenti e catalizzano la decarbossilazione degli aminoacidi nelle poliammine corrispondente in batteri e più alti organismi18,19 , 20 , 21 , 22. la mono – diidrossi composti (Figura 3) 4–7, 10–11 corrispondono ai idrossilati cadaverina, diammina ottenuta mediante decarbossilazione della L-lisina. La cadaverina è un elemento chiave per l’industria chimica, in particolare è un componente di polimeri in poliammide e poliuretano. Di conseguenza, produzione biologica di questo diammina da risorse rinnovabili ha attirato l’attenzione come un’alternativa al percorso a base di petrolio, e vari microrganismi sono stati progettati per questo scopo. In queste vie metaboliche, lisina DC (LDC) è l’enzima chiave. LDC è un enzima di PLP-dipendenti appartenenti al alanina racemasi (AR) strutturali famiglia23. I controller di dominio di PLP-dipendenti (PLP-DCs) sono noti per essere altamente substrato specifico. Tuttavia, alcuni enzimi proprio la capacità di promiscuità leggera, essendo attivo verso gli aminoacidi L-ornitina e la L-lisina, come ad esempio la LDC da Selenomonas rumirantium (LDCSrum), che ha simili costanti cinetiche per lisina e ornitina decarbossilazione24,25. Questo substrato esteso specificità rende questo enzima un buon candidato per la decarbossilazione di mono – e di-idrossi-L-lisina. Inoltre, per trovare DCs attivo verso i derivati dell’idrossile di lisina, abbiamo esaminato il contesto genomico dei geni che codificano gli enzimi αKAO. Infatti, nei genomi procariotici i geni che codificano per enzimi coinvolti nella via biosintetica del stessa generalmente sono co-localizzati in cluster genici. Il gene KDO2 (da Chitinophaga pinensis) è stato trovato co-localizzato con un gene che codifica per presunti PLP-DC (Figura 2). Al contrario, nessun gene che codifica per la DC è stato trovato quando si analizza il contesto genomico di dioxygenase del KDO1. La proteina PLP-DC da c. pinensis (DCCpin) pertanto è stata selezionata come candidato promising per catalizzare il passo di decarbossilazione della reazione cascade.
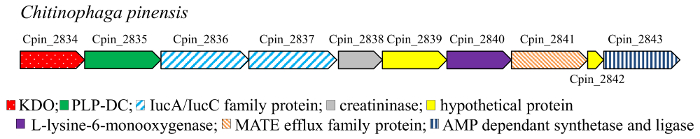
Figura 2: contesto Genomic del gene KDO2 in c. pinensis. Clicca qui per visualizzare una versione più grande di questa figura.
Di conseguenza, abbiamo progettato reazioni cascata enzimatica che coinvolgono diossigenasi e DCs per realizzare la sintesi di alcoli alifatici chirali β e γ-amminico da amminoacidi (Figura 3). Come precedentemente segnalato, l’ossidazione di C-H catalizzata dalla αKAO introduce il centro stereogenico idrossi-sostituiti con totale diastereoselettività; la chiralità Cβ/γ verrà mantenuta nel passaggio decarboxylative, che colpisce solo il carbonio Cα della molecola dell’amminoacido16.
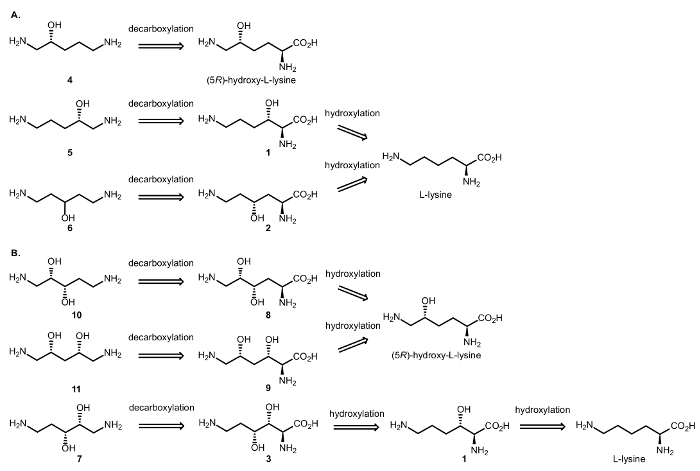
Figura 3: analisi retrosintetica. (A) Retrosynthesis di β e γ-amino alcoli (R) – 1,5 – diaminopentan-2-olo (4) da (5R) – idrossi – L-lisina e diaminopentan (S) – 1,5–2-ol (5) e 1,5-diaminopentan-3-ol (6) da L-lisina. (B) Retrosynthesis di β, γ e β, δ-amino dioli (2S, 3S) – 1,5 – diaminopentane-2,3-diolo (10) e (2R, 4S) – 1,5 – diaminopentane-2,4-diolo (11) a partire da (5R)- idrossi-L-lisina e (2R, 3R) – 1,5 – diaminopentane-2,3-diolo (7) a partire da L-lisina. Clicca qui per visualizzare una versione più grande di questa figura.
A partire da L-lisina e i suoi (5R)-derivato idrossilato, qui segnaliamo un due/tre step, una pentola, procedura enzimatica che unisce diossigenasi e PLP-DCs per ottenere l’obiettivo amino alcoli. Prima la sintesi a scala di laboratorio delle molecole bersaglio, il metodo è stato sviluppato presso la scala analitica per regolare le condizioni di reazione, ad esempio, le concentrazioni di enzimi, necessari per consentire la conversione completa dei materiali di partenza; Presentiamo questa procedura pure.
