We beschrijven een totale interne reflectie fluorescentie-nummer en helderheid (TIRF-N & B) Imaging benadering voor het bepalen van de gemiddelde oligomere toestand van receptor moleculen op het plasma membraan van levende cellen, gericht op het koppelen van de receptor assemblage dynamiek aan de biologische functie van de eiwitten (Figuur 1).
Bij extracellulaire ligand binding initiëren receptoren de intracellulaire signaaltransductie afhankelijk van hun conformatie, oligomerisatie, potentiële co-receptoren en membraan samenstelling. Ondanks het belang en de alomtegenwoordigheid van de receptor oligomerisatie, herkend als een belangrijke gebeurtenis in cellulaire signalering1,2,3,4,5,6, 7, enkele methoden kunnen clustering gebeurtenissen detecteren en meten van de mate van clustering experimentaal8,9. Het confocale volume (x, y ≈ 300 nm, z ≈ 900 nm) is onvoldoende verdwenen voor het aantonen van moleculaire interactie en stoichiometrie, zelfs na optimalisatie door beeld restauratie algoritmen10. De samenstelling van de subeenheid van eiwit oligomeren kan niet op zuiver ruimtelijke basis worden opgelost, zelfs niet door super-resolutie methoden bij x, y-resolutie van 20-70 nm zoals PALM11, Storm12en STED13. Bovendien kan de tijdelijke resolutie (in de volgorde van minuten per afbeelding) kinetiek niet in het bereik van seconden volgen. Enkelvoudige molecuul stap-bleaching lost de stoichiometrie van eiwit oligomeren alleen op als ze immobiel zijn14.
Een van de meest veelzijdige methoden om dichtheid en oligomisatie van fluorescently gelabelde eiwitten binnen enkele afbeeldingen te meten, is de analyse van ruimtelijke intensiteits verdeling (SpIDA), die afhankelijk is van ruimtelijke bemonstering. Het is toepasbaar op zowel chemisch vaste als levende cellen, en maakt het mogelijk de analyse van verschillende gebieden van belang van de cel tegelijkertijd met behulp van standaard fluorescentiemicroscopie15. Als alternatief zijn moment methoden, zoals fluorescentie-correlatie spectroscopie (FCS)16, foton Counting histogram (PCH)17, en aantal en helderheid (N & B)18,19, geschikt voor kwantitatief oligomeren Metingen. Deze methoden analyseren de intensiteit van de fluorescentie schommelingen die in de tijd kunnen worden waargenomen wanneer de fluor Foren zich in en uit een verlichtings volume verspreiden. De amplitudes van de intensiteits schommelingen kunnen op unieke wijze worden beschreven door de moleculaire helderheid van de de (ε) en het gemiddelde aantal fluor Foren (n) binnen het verlichtings volume17 (Figuur 2). Normaalgesproken kan de diffusiecoëfficiënt van de fluor Foren en het gemiddelde aantal moleculen (omgekeerd gerelateerd aan de G (0)-waarde) binnen het verlichtings volume worden verkregen door FCS20. Aangezien de diffusie tijd echter alleen wordt geschaald met de kubieke wortel van de massa, is FCS niet voldoende gevoelig voor het detecteren van veranderingen in molecuulmassa21. In de praktijk, single Color FCS kan niet detecteren door dimerisatie van membraan receptoren. PCH lost mengsels van verschillende oligomeren nauwkeurig op. Met behulp van meer dan twee momenten van de amplitude-verdeling, detecteert het moleculen van verschillende helderheid die hetzelfde verlichtings volume innemen. Scanning FCS22 en ontwikkelingen, zoals de interessante pair-correlatie van moleculaire helderheid (pcomb) approach23, geïntroduceerd om het bereik van toepasbaarheid van fluorescentie correlatie methoden in biologische systemen uit te breiden24 , blijven methoden met één punt ontbreken van snelle metingen in een groot gebied van een cel, waarbij veel opeenvolgende waarnemingen bij elke pixel en het verzamelen van gegevens in de volgorde van seconden worden vereist.
N & B is een vereenvoudigde versie van PCH die alleen de eerste en tweede momenten van de amplitude van de fluorescentie verdeling beschouwt, namelijk de gemiddelde intensiteit, , en de variantie, σ2 (Figuur 2)18,19 en daarom kan het de molaire fractie van onbekende oligomeren in een mengsel niet bepalen, maar slechts schattingen van de gemiddelde oligomerisatie toestand van het mengsel. Niettemin heeft N & B het voordeel van het werken met relatief kleinere tijdreeksen beelden van levende cellen dan PCH per pixel, simpelweg door de schommelingen op de tijd van de intensiteit van de fluorescentie te bewaken. Omdat N & B de tijd per pixel tot een paar microseconden verlaagt, kan het snelle oligomerisatie kinetiek volgen op grote celgebieden, waardoor beeld verwerving op een tijdschaal van seconden in raster scanning microscopie (bijv. confocale, 2-photon) en milliseconden op camera gebaseerde microscopie (bijv. TIRFM).
Verschillende rapporten hebben aangetoond dat N & B het aantal subeenheden in eiwit clusters kwantificeren door uitgebreide celregio’s. Paxillin-EGFP-clusters werden aangetroffen op de adhesieve plekken in de CHO-K1-cellen25, en de intracellulaire aggregatie van het pathogene Httex1p peptide werd beschreven in cos-7-cellen26. N & B werd toegepast voor het volgen van de ligand-gestuurde oligomisatie van de ErbB-receptor27, en het effect van de LIGAND FGF21 op Klothob (klb) en FGFR1c in HeLa-cellen28. De combinatie van tirf-beeldvorming en N & B-analyse werd gebruikt om aan te tonen dat dynamin-2 voornamelijk tetramere is in het gehele celmembraan29. We hebben N & B toegepast op zowel raster scanning als tirf-beelden om ligand-gestuurde door dimerisatie van upar en FGFR1 celmembraan receptoren30,31te bewijzen.
Fluorescentie-correlatie methoden, zoals N & B, FCS en PCH, zijn gebaseerd op de gedachte dat in een open volume het bezettings getal van deeltjes een Poisson-verdeling volgt. Omdat alleen de fotonen die de fluoroforen uitzenden kunnen worden gedetecteerd, de gemiddelde waarde voor een gemeten fluorescentie intensiteit versus tijd in een pixel van het beeld,  , is het product van het gemiddelde aantal fluor Foren in het verlichtings volume, n, en hun moleculaire helderheid, ε17:
, is het product van het gemiddelde aantal fluor Foren in het verlichtings volume, n, en hun moleculaire helderheid, ε17:

waarbij ε wordt uitgedrukt als het aantal fotonen dat per tijdseenheid (conventioneel per seconde) per molecuul wordt uitgezonden wanneer het molecuul zich in het midden van het verlichtings volume bevindt.
Helderheid is een eigenschap van elke de in een bepaalde overname instellen, terwijl intensiteit de som is van alle bijdragen van alle fluor Foren. Bij biologische wedstrijden zal de helderheid toenemen met de toename van het aantal fluoroforen dat samen fluctueert, wat informatie geeft over de oligomerisatie toestand van het fluorescently-gelabelde eiwit. De fluctuatie amplituden op een bepaalde pixel wordt gemeten aan de variantie van het fluorescentie signaal, σ2:

Wanneer het gemiddelde van het kwadraat van de intensiteit  , en het kwadraat van het gemiddelde van de intensiteit
, en het kwadraat van het gemiddelde van de intensiteit  , worden berekend op basis van de individuele intensiteitswaarden in elke pixel van elk frame:
, worden berekend op basis van de individuele intensiteitswaarden in elke pixel van elk frame:
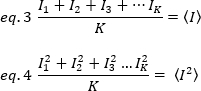
waarbij K het totale aantal frames in de tijdreeks is. Experimenteel is het noodzakelijk om voor de hele Afbeeldingsreeks de variantie te berekenen die de spreiding van de individuele intensiteitswaarden op elke pixel van één afbeelding rond de gemiddelde intensiteitswaarde beschrijft. De variantie omvat alle schommelingen van verschillende oorsprongen. Bij een eerste benadering kan de variantie als gevolg van de diffusisolerende deeltjes in het verlichtings volume σ20worden gescheiden van de variantie als gevolg van de detector schot ruis, σ2d. De twee varianties zijn onafhankelijk; Zo wordt de totale variantie gegeven door hun som:

De variantie, als gevolg van moleculaire schommelingen in en uit het detectie volume, is lineair afhankelijk van de moleculaire helderheid en intensiteit:

EQ. 6 opnieuw rangschikken volgens EQ. 1:

Volgens het typische concept in fluorescentie correlatie spectroscopie, stelt vergelijking 7 dat de variantie als gevolg van het aantal schommelingen afhangt van het kwadraat van de deeltjes helderheid.
Vervolgens is de variantie als gevolg van schommelingen in de detector een lineaire functie van de gedetecteerde intensiteit, onder de aanname dat de detector onder de verzadigings limiet19wordt bediend:

In het geval van foon tellings detectoren a= 1 en c= 0 is de variantie van de detector gelijk aan de gemiddelde intensiteit:

Om deze concepten op echte metingen in levende cellen toe te passen, definiëren Gratton en collega’s18 de schijnbare helderheid, B, voor elke pixel als de verhouding van de variantie over de gemiddelde intensiteit:

B is de parameter die experimenteel wordt gemeten. In dit werk, time series beelden van FGFR1 receptoren op het plasma membraan van HeLa cellen worden gevangen door TIRF microscopie en de gemiddelde schijnbare helderheid, B, wordt bepaald door de N & B analyse. Vervolgens, na toevoeging van FGF2, opeenvolgende tijdreeksen worden gevangen om de veranderingen in de zelf-assemblage van de receptor moleculen in het membraanoppervlak na stimulatie van de receptor met de canonieke ligand te volgen.
Aangezien de detector van de TIRF-Microscoop echter een EMCCD-camera is, moet de uitdrukking voor de schijnbare helderheid worden gewijzigd als19:

waarbij Offset de intensiteit offset is van de detectie-elektronica die kenmerkend is voor de detector instellingen. De variantie en gemiddelde intensiteit voor een analoge detector worden respectievelijk gegeven door:
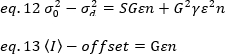
waarbij G de analoge winst in digitale niveaus (DL/fotonen), S, de digitale niveaus per foton19, wordt gegeven door de helling van een intensiteit versus variantie plot voor een lichtbron met constante intensiteit (geen tijdelijke schommelingen). De γ-factor is gerelateerd aan de vorm van het pixel detectie volume. Volgens Hassler et al.32is de γ-factor gelijk aan 0,3 voor tirf-beeldvorming die werkt op de maximale Gain van de detectie camera19. De parameters offset, S en G zijn kenmerken van de camera en de Microscoop. De schijnbare helderheid, B, wordt verkregen door het herschikken van EQ. 11 volgens EQ. 12 en 13:

Experimenteel is ε een complexe functie van laserintensiteit en de detectie-efficiëntie van het systeem. Niettemin, aangezien B/S lineair afhankelijk is van ε, is het alleen belangrijk om de relatieve waarde van ε te bepalen voor een bepaalde detectiemodus:

waar ε ‘ evenredig is aan ε. Toch wordt een kalibratie uitgevoerd met een interne referentie.
