Canlı hücrelerin plazma zarındaki reseptör moleküllerinin ortalama oligomerik durumunu belirlemek için toplam Internal Yansıma Floresan-Sayı ve Parlaklık (TIRF-Sayı ve Parlaklık) görüntüleme yaklaşımını, reseptör diziliyi bağlamayı amaçlayan bir toplam iç yansıma floresan-sayı ve parlaklık (TIRF-N&B) görüntüleme yaklaşımını tanımladık. proteinlerin biyolojik işlevine dinamikler (Şekil 1).
Hücre dışı ligand bağlanması üzerine reseptörler konformasyonlarına, oliomerizasyonlarına, potansiyel ko-reseptörlerine ve membran kompozisyonlarına bağlı olarak hücre içi sinyal iletimini başlatırlar. Reseptör oligomerizasyonunun önemi ve her yerde, hücreselsinyalizasyon daönemli bir olay olarak kabul rağmen 1,2,3,4,5,6, 7, birkaç yöntem kümeleme olayları algılayabilir ve deneyselkümelemederecesini ölçmek 8,9. Konfokal hacim (x,y 』 300 nm, z 』 900 nm) moleküler etkileşimi ve stokiyometriyi kanıtlamak için yeterli olarak çözülmüş, görüntü restorasyon algoritmaları tarafından optimizasyon dan sonra bile10. Protein oligomerlerinin alt birim bileşimi, PALM11, STORM12ve STED13gibi 20-70 nm x,y çözünürlükte süper çözünürlük yöntemleriyle bile tamamen mekansal olarak çözülemez. Ayrıca, onların zamansal çözünürlük (görüntü başına dakika sırasına göre) saniye aralığında kinetik takip edemez. Tek moleküllü adım ağartma protein oligomerlerinin stoiyometrisini ancak hareketsiz olduklarında çözer14.
Floresan etiketli proteinlerin tek bir görüntü içinde yoğunluk ve oliomerizasyonunu ölçmek için en çok yönlü yöntemlerden biri, mekansal örneklemeye dayanan mekansal yoğunluk dağılım analizidir (SpIDA). Hem kimyasal olarak sabit hem de canlı hücreler için geçerlidir ve aynı anda standart floresan mikroskobu15kullanarak hücrenin ilgi çeşitli bölgelerin analizisağlar. Alternatif olarak, floresan-korelasyon spektroskopisi (FCS)16,foton sayma histogramı (PCH)17ve Sayı ve Parlaklık (N&B)18,19gibi moment yöntemleri kantitatif oligomer için uygundur Ölçüm. Bu yöntemler, floresan ların aydınlatma hacmine girip çıktığı zaman gözlemlenebilen floresan yoğunluk dalgalanmalarını analiz eder. Yoğunluk dalgalanmalarının genlikleri, floroforun moleküler parlaklığı (ε) ve aydınlatma hacmindeki ortalama florofor (n) sayısı(şekil 2)ile benzersiz bir şekilde tanımlanabilir. Tipik olarak, floroforların difüzyon katsayısı ve aydınlatma hacmi içindeki ortalama molekül sayısı (G(0) değeriile ters orantılı) FCS20ile elde edilebilir. Ancak, difüzyon süresi sadece kütlenin kübik kökü ile ölçeklenir beri, FCS moleküler kütle değişiklikleri tespit etmek için yeterince duyarlı değildir21. Uygulamada, tek renkli FCS membran reseptörlerinin dimerizasyon unu algılayamaz. PCH, farklı oligomerlerin karışımlarını doğru bir şekilde çözer. Genlik dağılımının iki dakikadan fazlaını kullanarak, aynı aydınlatma hacmini kaplayan farklı parlaklıktaki molekülleri algılar. Tarama FCS22 ve gelişmeler, moleküler parlaklık ilginç çift korelasyon gibi (pCOMB) yaklaşım23, biyolojik sistemlerde floresan korelasyon yöntemlerinin uygulanabilirlik aralığını genişletmek için tanıtıldı24 , bir hücrenin geniş bir alanında hızlı ölçüm yeteneği nden yoksun tek noktalı yöntemler kalır, saniye sırasına göre her piksel ve veri toplama birçok ardışık gözlemler gerektiren.
N&B, PCH’nin floresan dağılımının genliğin inkişanın sadece birinci ve ikinci anlarını, yani ortalama yoğunluğunu, , ve varyans, σ2 (Şekil 2) 18,19’udikkate alan basitleştirilmiş bir sürümüdür. ve bu nedenle, bir karışımda bilinmeyen oligomerlerin molar fraksiyonunu belirleyemez, ancak sadece karışımın ortalama oliomerizasyon durumunu tahmin eder. Bununla birlikte, N & B sadece floresan yoğunluğu zamanında dalgalanmaları izleyerek, bir piksel-piksel bazında PCH daha canlı hücrelerin görüntüleri nispeten daha küçük zaman serisi ile çalışma avantajı vardır. N&B piksel başına zamanı birkaç mikrosaniyeye düşürebildiği için, büyük hücre alanları üzerinde hızlı oligomerizasyon kinetiği izleyebilir ve raster tarama mikroskopisinde (örn. konfokal, 2-foton) ve milisaniyelerde saniyeler ölçeğinde görüntü elde edilmesine izin verir. kamera tabanlı mikroskopide (örn. TIRFM).
Çeşitli raporlar, Genişletilmiş hücre bölgelerini görüntüleyerek protein kümelerinde alt birim sayısını ölçebilme yeteneğini göstermiştir. PAXIllin-EGFP kümeleri CHO-K1 hücrelerinde adezyon bölgelerinde tespit edildi25, ve patojenik Httex1p peptid in hücre içi agregasyonu COS-7 hücrelerinde tanımlanmıştır26. N & B ErbB reseptörünün ligand tahrikli oligomerizasyonu aşağıdaki için uygulandı27, ve HeLa hücrelerinde Klothob üzerinde ligand FGF21 etkisi (KLB) ve FGFR1c HeLa hücrelerinde28. TIRF görüntüleme ve N & B analizi kombinasyonu dinamin-2 öncelikle tüm hücre zarı29boyunca tetramerik olduğunu göstermek için kullanılmıştır. UPAR ve FGFR1 hücre zarı reseptörlerinin ligand tahrikli dimerizasyonunu kanıtlamak için hem raster tarama sınahem de TIRF görüntülerine N&B uyguladık30,31.
N&B, FCS ve PCH gibi floresan korelasyon yöntemleri, açık bir hacimde parçacıkların işgal sayısının Bir Poisson dağılımını izlediği fikrine dayanır. Sadece floroforların emit tespit edilebildiği fotonlar, görüntünün  bir pikselinde ölçülen floresan yoğunluğuile zaman arasındaki ortalama değer, aydınlatma hacmindeki ortalama floresan sayısının, n ve bunların moleküler parlaklık, ε17:
bir pikselinde ölçülen floresan yoğunluğuile zaman arasındaki ortalama değer, aydınlatma hacmindeki ortalama floresan sayısının, n ve bunların moleküler parlaklık, ε17:

molekül aydınlatma hacminin merkezindeyken molekül başına zaman birimi başına yayılan foton sayısı (geleneksel olarak saniyede) olarak ifade edilir.
Parlaklık belirli bir edinim kurmak her florofor bir özelliğidir, yoğunluk tüm floroforlardan tüm katkıların toplamı ise. Biyolojik yarışmalarda, birlikte dalgalanan florofor sayısının artmasıyla parlaklık artarak floresan etiketli proteinin oliomerizasyon durumu hakkında bilgi verecektir. Belirli bir pikseldeki dalgalanma genlikleri floresan sinyalinin varyansından ölçülür, σ2:

Yoğunluk  karesinin ortalaması ve yoğunluk ortalamasının karesi,
karesinin ortalaması ve yoğunluk ortalamasının karesi,  her karenin her pikselindeki tek tek yoğunluk değerlerinden hesaplanır:
her karenin her pikselindeki tek tek yoğunluk değerlerinden hesaplanır:
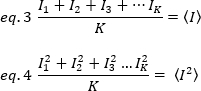
k zaman serisindeki toplam kare sayısıdır. Deneysel olarak, tüm görüntü serisi için ortalama yoğunluk değeri etrafında tek bir görüntünün her pikselinde tek tek yoğunluk değerlerinin dağılım açıklayan varyans hesaplamak için gereklidir. Varyans, farklı kökenlerdeki tüm dalgalanmaları içerir. İlk yaklaşık olarak, aydınlatma hacmindeki difüzyon parçacıklar nedeniyle varyans, σ20, dedektör atış gürültüsü nedeniyle varyans ayrılabilir, σ2d. İki varyans bağımsızdır; böylece, toplam varyans onların toplamı ile verilir:

Algılama hacmindeki ve dışında meydana gelen moleküler dalgalanmalar nedeniyle varyans, moleküler parlaklığa ve yoğunluğa doğrusal olarak bağlıdır:

Eq 1’e göre eq 6’nın yeniden düzenlenmesi:

Floresan korelasyon spektroskopisinde tipik kavrama göre, 7.
Daha sonra, dedektör dalgalanmaları nedeniyle varyans dedektör doygunluk sınırı nın altında çalıştırılır varsayımı altında, tespit yoğunluğunun doğrusal bir fonksiyonudur19:

Foton sayma dedektörleri durumunda a=1 ve c=0, böylece dedektör varyans ortalama yoğunluğuna eşittir:

Bu kavramları canlı hücrelerdeki gerçek ölçümlere uygulamak için Gratton ve meslektaşları18, her piksel için görünür parlaklığı, B’yi ortalama yoğunluk üzerinde varyans oranı olarak tanımlar:

B, deneysel olarak ölçülen parametredir. Bu çalışmada, HeLa hücrelerinin plazma zarındaki FGFR1 reseptörlerinin zaman serisi görüntüleri TIRF mikroskobu ile yakalanır ve ortalama parlaklık olan B, N&B analizi ile belirlenir. Daha sonra, FGF2 eklenmesinden sonra, ardışık zaman serileri kanonik ligand ile reseptör uyarılmasından sonra membran yüzeyinde reseptör moleküllerinin kendi kendine montaj değişiklikleri takip yakalanır.
Ancak, TIRF mikroskobu dedektörü bir EMCCD kamera olduğundan, görünür parlaklık için ifade19olarak değiştirilmesi gerekir:

ofset dedektör ayarlarının bir özelliğidir algılama elektroniği yoğunluğu ofset nerede. Bir analog dedektör için varyans ve ortalama yoğunluğu sırasıyla tarafından verilir:
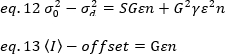
nerede G dijital düzeylerde analog kazanç (DL / foton), S, foton başına dijital düzeyleri19, sabit yoğunluğu (hiçbir zamansal dalgalanmalar) ile bir ışık kaynağı için bir yoğunluk karşı varyans arsa eğimi tarafından verilir. Γ faktörü piksel algılama hacminin şekli ile ilgilidir. Hassler ve ark.32’yegöre γ faktörü, algılama kamerası19’unmaksimum kazancıyla çalışan TIRF görüntülemeiçin 0,3’e eşittir. Ofset, S ve G parametreleri kamera ve mikroskop özellikleridir. Görünür parlaklık, B, eq 11 eq 12 ve 13 göre yeniden düzenlenmesi ile elde edilir:

Deneysel olarak, ε lazer yoğunluğu ve sistemin algılama verimliliği karmaşık bir fonksiyonudur. Bununla birlikte, B/S doğrusal olarak ε’ya bağlı olduğundan, yalnızca belirli bir algılama modu için ε’nin göreli değerini belirlemek önemlidir:

ε’ ile orantılı dır. Yine de, bir kalibrasyon dahili bir başvuru kullanılarak gerçekleştirilir.
