Los latidos del corazón en las sanguijuelas son impulsados por una GPC que consiste en 9 pares bilaterales de interneuronas cardíacas (HN) distribuidas en la mayor cantidad de ganglios segmentarios de la mitad del cuerpo. En el núcleo de la GPC se encuentran pares mutuamente inhibitorios de interneuronas ubicadas en los ganglios segmentarios3º y4º que forman osciladores semicéntricos (HCO)(Figura 1A). Estas neuronas continúan estallando cuando se aíslan sinápticamente farmacológicamente usando bicucullina1. Otros, como el par en los ganglios segmentarios7 (el foco de este protocolo), también son ráfagas, capaces de producir actividad de estallido cuando se aíslan sinápticamente. No están mutuamente conectados y reciben solo entrada descendente, y por lo tanto se aíslan fácilmente al separar el ganglio del resto del cordón nervioso. Esta actividad de estallido independiente es sensible a la corriente de fuga introducida causada por la penetración con microelectrodos afilados para la grabación, pero estalla vigorosamente cuando se graba con métodos de parche suelto1.
Se han modelado tanto las neuronas HN individuales como las HN HNO HCOs (modelos de compartimento isopotencial único basados en Hodgkin-Huxley de neuronas HN que contienen todas las corrientes sinápticas y de voltaje identificadas experimentalmente), y todas las características de estallido del sistema vivo se han capturado con éxito2. La miomodulina, un neuropéptido endógeno en las sanguijuelas, disminuye notablemente el período (T) del ritmo de estallido de las neuronas HN aisladas y las HN HNo. Este modulador actúa para aumentar la corriente h (corriente interna activada por hiperpolarización, Ih)y para disminuir labombaI3. Esta observación llevó a la exploración de cómo labomba I interactúa con Ih,y cómo su co-modulación contribuye a la actividad rítmica de las neuronas HN. La activación de la bomba mediante el aumento de [Na+]i (utilizando el ionóforo monensina) acelera el ritmo de estallido de HN tanto en HN HCOs como en neuronas HN aisladas4. Esta aceleración dependía de Ih. Cuando Ih estaba bloqueado (2 mM Cs+),el período de ráfaga no se alteraba por este método de activación de la bomba; sin embargo, la duración de la ráfaga (BD) se redujo y el intervalo de interbursura (IBI) aumentó tanto en HN HCOs como en neuronas HN aisladas4.
Para este protocolo, todas las corrientes de una neurona HN(7) viva, incluida la corriente de bomba, I bomba,se incorporan en el modelo HN de la siguiente manera:
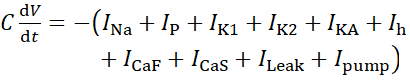 (1)
(1)
donde C es la capacitancia de membrana (en nF), V es el potencial de membrana (en V), t es el tiempo (en s). Las descripciones y ecuaciones detalladas de la corriente iónica se han descrito en otra parte2,4. La neurona modelo HN completa se ejecuta en tiempo real (Figura 2). El software estará disponible en GitHub en el momento de su publicación y será adecuado para ejecutarse en la placa de procesamiento de señal digital descrita en la Tabla de materiales. Aquí, el foco de investigación es la corriente de la bomba Na+/ K+ (bombaI) y las corrientes de voltaje que contribuyen con un flujo significativo de Na+: una corriente rápida de Na+ (INa) y una corriente persistente de Na+ (IP). Las conductancias máximas de estas corrientes son 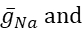
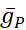 respectivamente. La bomba Na+/K+ intercambia tres iones intracelulares Na+ por dos iones K+ extracelulares, produciendo así una corriente neta hacia el exterior. Es importante destacar que bombea 3 veces más Na+ fuera de la neurona de lo que indica esta corriente, lo cual es importante para calcular la concentración intracelular de Na+.
respectivamente. La bomba Na+/K+ intercambia tres iones intracelulares Na+ por dos iones K+ extracelulares, produciendo así una corriente neta hacia el exterior. Es importante destacar que bombea 3 veces más Na+ fuera de la neurona de lo que indica esta corriente, lo cual es importante para calcular la concentración intracelular de Na+.
La corriente de bomba Na+/K+ depende de las concentraciones intracelulares de Na+ y se expresa mediante la siguiente función sigmoidal:
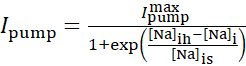 (2)
(2)
donde [Na]i es la concentración intracelular de Na+, 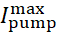 es la corriente máxima de la bomba Na+/K+, [Na]ih es la concentración intracelular de Na+ para la media activación de la bomba Na+/K+, y [Na]es la sensibilidad de la bomba Na+/K+ a [Na]i. [Na]i se construye como resultado de las afluencias de Na+ transportadas por IP e INa y se ve disminuida por el eflujo na+ de la bomba Na+/ K+. La contribución de Ih e ILeak al flujo total deNa+ es pequeña y no se considera en el modelo en tiempo real.
es la corriente máxima de la bomba Na+/K+, [Na]ih es la concentración intracelular de Na+ para la media activación de la bomba Na+/K+, y [Na]es la sensibilidad de la bomba Na+/K+ a [Na]i. [Na]i se construye como resultado de las afluencias de Na+ transportadas por IP e INa y se ve disminuida por el eflujo na+ de la bomba Na+/ K+. La contribución de Ih e ILeak al flujo total deNa+ es pequeña y no se considera en el modelo en tiempo real.
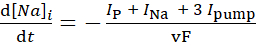 (3)
(3)
donde, v es el volumen (~6.7 pL) del reservorio intracelular de Na+, F es la constante de Faraday, y la concentración extracelular de Na+ se mantiene constante.
Se han diferenciado las conductancias dependientes de voltaje y fugas, que responden al potencial de membrana, de la corriente de la bomba, que está regulada por la concentración intracelular calculada de Na+ ([Na+]i). [Na+] i se construye a través de na+ entrada a través de la corriente rápida Na+ (INa) que produce potenciales de acción (picos) y la corriente persistente Na+ (IP) que proporciona la despolarización para soportar el aumento. [Na+] i es, a su vez, reducido por la acción de la bomba a través de la extrusión deNa+. Se han asumido valores basales de HN vivo de 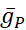 (5nS) y
(5nS) y 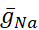 (150 nS), y tenemos en cuenta cualquier abrazadera dinámica añadida.
(150 nS), y tenemos en cuenta cualquier abrazadera dinámica añadida.
El objetivo del protocolo descrito aquí es manipular la bomba de forma precisa y reversible en tiempo real para descubrir cómo interactúa con las corrientes de voltaje (corriente persistente de Na+ en el protocolo actual) para controlar el estallido rítmico en HNs individuales. Para lograr este objetivo, se utilizó una abrazadera dinámica, que introduce artificialmente, por orden, una cantidad precisa de cualquier corriente que se puede calcular a medida que se ejecuta el modelo. Este método tiene ventajas sobre la manipulación farmacológica de la bomba, que afecta a todo el tejido, puede tener efectos fuera del objetivo que a menudo son difíciles de revertir y no se pueden manipular con precisión. La abrazadera dinámica5,6 lee el voltaje de una neurona registrada en tiempo real(Figura 1B)y calcula e inyecta, en tiempo real, la cantidad de cualquier corriente basada en ecuaciones del modelo y los valores establecidos de 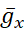 cualquiera o
cualquiera o 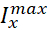 . Métodos similares se pueden aplicar fácilmente a cualquier neurona que se pueda registrar intracelularmente. Sin embargo, los parámetros tendrán que ser reescalados a la neurona elegida, y la neurona debe aislarse de las entradas sinápticas, por ejemplo, farmacológicamente.
. Métodos similares se pueden aplicar fácilmente a cualquier neurona que se pueda registrar intracelularmente. Sin embargo, los parámetros tendrán que ser reescalados a la neurona elegida, y la neurona debe aislarse de las entradas sinápticas, por ejemplo, farmacológicamente.
