De productie van de meeste eukaryote boodschapper-RNA’s (mRNA’s) omvat verwijdering van introns en ligatie van exonen in een nucleair proces dat pre-mRNA-splicing wordt genoemd1. Twee klassen van RNA-eiwitcomplexen (RRP’s) sturen de verwerking van pre-messenger RNA tot volwassen mRNA via spliceosomale complexen. Eén klasse, ontluikende pre-messenger RRP’s, wordt co-transcriptioneel gevormd door de binding van heterogene nucleaire RNP-eiwitten en andere RNA-bindende eiwitten, waaronder enkele leden van de SR-familie, die hnRNP-complexen opleveren2. De tweede klasse, uracil-rijke kleine nucleaire RRP’s (U snRNPs met U1, U2, U4, U5 en U6 snRNAs) is geassocieerd met U-specifieke en kerneiwitten3,4. De U snRNPs interageren op een geordende manier met specifieke regio’s van pre-messenger RRP’s in een dynamische remodelleringsroute, terwijl introns worden weggesneden en exonen worden geligeerd om volwassen mRNPs5 te produceren. Veel extra nucleaire eiwitten nemen deel aan deze verwerkingsgebeurtenissen6.
Galectine-1 (Gal1) en galectine-3 (Gal3) zijn twee eiwitten die vereiste factoren zijn in de splicingroute, zoals aangetoond door depletie-reconstitutiestudies7,8. Verwijdering van beide galectines uit het splitsen van competente nucleaire extracten (NE) elimineert spliceosoomassemblage en splicingactiviteit in een vroeg stadium. Toevoeging van beide galectine aan zo’n dubbel uitgeput NE herstelt beide activiteiten. Gal1 en Gal3 zijn componenten van actieve spliceosomen zoals blijkt uit specifieke immunoprecipitatie van pre-mRNA, splicing intermediates en volwassen mRNA door antiserum specifiek voor Gal1 of Gal39. Belangrijk is dat Gal3 zich associeert met endogene U-snRNA die deeltjes in het NE bevatten buiten de splicingroute, zoals blijkt uit de precipitatie van snRNPs door anti-Gal3 antisera10. Ten slotte verandert het uitschakelen van Gal3 in HeLa-cellen de splicingpatronen van talrijke genen11.
In NE voorgeïncubeerd om voorgevormde spliceosomen12 te demonteren, worden snRNPs gevonden in meerdere complexen die sedimenteren in glycerolgradiënten van 7S tot groter dan 60S. Hoewel glycerolgradiëntfractionering een veelgebruikte techniek is voor de isolatie van spliceosomale complexen en componenten (zie bijvoorbeeld referenties13,14,15), hebben we deze methode uitgebreid door specifieke fracties te karakteriseren met behulp van antilichaamimmunoprecipitaties. Een snRNP-sedimentering op 10S bevat alleen U1-snRNA samen met Gal3. Immunoprecipitatie van de 10S-fractie met antisera specifiek voor Gal3 of U1 snRNP co-precipiteert zowel U1 als Gal3, wat aangeeft dat sommige van de U1 snRNP-monodeeltjes gebonden zijn aan Gal310. Aangezien U1 snRNP het eerste complex is dat bindt aan pre-mRNP in spliceosomale assemblage1,5, vertegenwoordigt deze stap een potentiële ingangsplaats voor Gal3 in de splicingroute. Op basis hiervan toonden we aan dat 10S Gal3-U1 snRNP monodeeltjes gebonden aan anti-Gal3 bevattende kralen de splicingactiviteit herstelden tot een U1 snRNP uitgeput NE, waardoor dit complex werd vastgesteld als één mechanisme waarmee Gal3 wordt gerekruteerd in de spliceosomale route16. Dit in tegenstelling tot pogingen om spliceosomen te isoleren in specifieke stadia van de splicingreactie en het catalogiseren van de geassocieerde factoren17,18. In dergelijke studies wordt de aanwezigheid van bepaalde factoren op een bepaald moment vastgesteld, maar niet het mechanisme waarmee ze werden geladen.
We hadden eerder in detail de bereiding van NE, het splicing substraat, de assemblage van het splicing reactiemengsel en de analyse van producten beschreven in onze documentatie van de rol van galectinen in pre-mRNA splicing19. We beschrijven nu de experimentele procedures voor fractionering van nucleaire extracten om een fractie verrijkt in Gal3 – U1 snRNP-complex te verkrijgen en voor immunoselectie van het laatste complex om splicingactiviteit te reconstitueren in een U1-uitgeput nucleair extract.
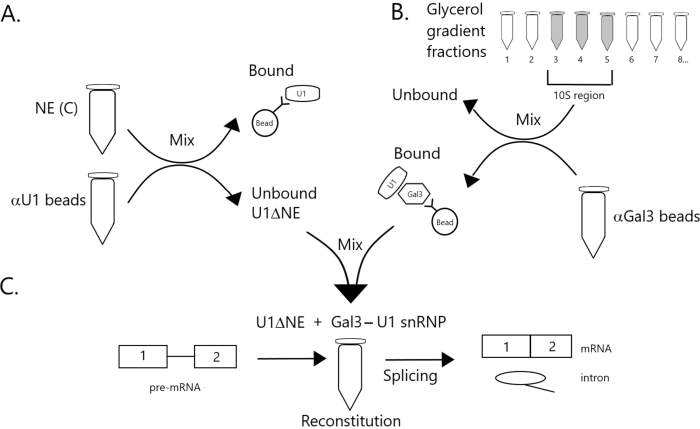
Figuur 1: Schematisch diagram ter illustratie van de complementatie van splicingactiviteit in nucleair extract uitgeput van U1 snRNP door een Gal3-U1 snRNP-complex op kralen. (A) NE in Buffer C (NE(C)) wordt geïncubeerd met eiwit A-Sepharose kralen covalent gekoppeld aan anti-U1 snRNP (αU1 kralen). De ongebonden fractie is uitgeput van U1 snRNP (U1ΔNE). (B) NE in Buffer D (NE(D)) wordt gefractioneerd over een glycerolgradiënt van 12%-32% door ultracentrifugatie. Fracties die overeenkomen met het 10S-gebied (fracties 3-5) worden gecombineerd en gemengd met kralen die covalent zijn gekoppeld aan anti-Gal3-antilichamen (αGal3-kralen). Het materiaal gebonden aan de kralen bevat een Gal3-U1 snRNP monodeeltje. (C) Het Gal3-U1 snRNP-complex uit deel (B) wordt gemengd met U1ΔNE uit deel (A) in een splicing-assay met behulp van 32P-gelabeld MINX pre-mRNA-substraat en de tussenproducten en producten van de splicingreactie worden geanalyseerd door gel-elektroforese en autoradiografie. Klik hier om een grotere versie van deze figuur te bekijken.
