הייצור של רוב RNAs שליח eukaryotic (mRNAs) כרוך בהסרת introns וקשירה של exons בתהליך גרעיני המכונה חיבור לפני mRNA1. שני סוגים של מתחמי חלבון RNA (RNPs) מכוונים את העיבוד של RNA לפני השליח ל- mRNA בוגר באמצעות מתחמים spliceosomal. מחלקה אחת, RNPs טרום שליח המתהווה, נוצרת שיתוף תמלול על ידי כריכה של חלבונים גרעיניים הטרוגניים וחלבונים אחרים מחייבים RNA, כולל כמה מבני משפחת SR, מניב מתחמי hnRNP2. ה-RNPs הגרעיניים הקטנים העשירים בארסיל (U snRNPs עם U1, U2, U4, U5 ו-U6 snRNAs) קשורים לחלבונים ספציפיים ל-U ולחלבוני ליבה3,4. ה- snRNPs של ארה”ב מתקשרים באופן מסודר עם אזורים ספציפיים של RNPs לפני השליח במסלול שיפוץ דינמי כאשר אינטרונים נקטעים ואקסונים קשורים כדי לייצר mRNPs5 בוגרים. חלבונים גרעיניים רבים נוספים משתתפים באירועי עיבוד אלה6.
גלקטין-1 (Gal1) וגלקטין-3 (Gal3) הם שני חלבונים הנדרשים במסלול ההשתלבות כפי שמוצג על ידי מחקרי דלדול-שיקום7,8. הסרת שני הגלקטינים מליזוב תמציות גרעיניות מוסמכות (NE) מבטלת את ההרכבה המשולבת ואת פעילות ההשתלבות בשלב מוקדם. תוספת של כל גלקטין ל- NE מרוקן כפליים משחזרת את שתי הפעילויות. Gal1 ו- Gal3 הם מרכיבים של spliceosomes פעיל כפי שמעידים אימונופרציפיטציה ספציפית של טרום mRNA, שילוב ביניים, ו- mRNA בוגר על ידי אנטי-ספירום ספציפי עבור Gal1 או Gal39. חשוב לציין, Gal3 מקשר עם snRNA U אנדוגני המכיל חלקיקים ב- NE מחוץ למסלול ההשתלבות כפי שמוצג על ידי משקעים של snRNPs על ידי אנטי-Gal3 antisera10. לבסוף, השתקה של Gal3 בתאי HeLa משנה דפוסי שכפול של גנים רבים11.
ב- NE דגירה מראש כדי לפרק spliceosomes preformed12, snRNPs נמצאים במתחמים מרובים המשקעים בשיפועים גליצריל מ 7S ליותר מ 60S. למרות שבר הדרגתי גליצול היא טכניקה נפוצה לבידוד של מתחמים ורכיבים spliceosomal (ראה הפניות13,14,15 למשל), הרחבנו שיטה זו על ידי אפיון שברים ספציפיים באמצעות חיסונים נוגדנים. משקעים snRNP ב- 10S מכיל רק U1 snRNA יחד עם Gal3. אימונופרציפיטציה של שבר 10S עם אנטי-רה ספציפית עבור Gal3 או U1 snRNP שיתוף מזרז הן U1 והן Gal3 המציין כמה חלקיקים U1 snRNP קשורים Gal310. מכיוון ש- U1 snRNP הוא המתחם הראשון שנקשר ל- pre-mRNP בהרכבה spliceosomal1,5, שלב זה מייצג אתר כניסה פוטנציאלי עבור Gal3 למסלול ההשתתפות. על בסיס זה, הראינו כי 10S Gal3-U1 snRNP מונו-חלקיקים הקשורים אנטי-Gal3 המכיל חרוזים משוחזרים פעילות splicing ל- U1 snRNP מרוקן NE, הקמת מורכב זה כמנגנון אחד שבאמצעותו Gal3 מגויס לתוך המסלול spliceosomal16. זאת בניגוד לניסיונות לבודד את התרסיסים בשלבים ספציפיים בתגובת ההשתלבות ולקטלג את הגורמים הקשורים 17,18. במחקרים כאלה, נוכחותם של גורמים מסוימים בנקודת זמן כלשהי מובהר אך לא המנגנון שבאמצעותו הם נטענו.
תיארנו בעבר בפירוט את הכנת NE, את מצע ההשתלבות, את ההרכבה של תערובת התגובה המשתלבת, ואת הניתוח של מוצרים בתיעוד שלנו של תפקיד הגלקטינים ב- splicing לפני mRNA19. כעת אנו מתארים את ההליכים הניסיוניים לפירוק תמציות גרעיניות כדי להשיג שבר מועשר במתחם Gal3 – U1 snRNP ולבחירה חיסונית של המתחם האחרון כדי לחדש את פעילות ההכלאה בתמצית גרעינית מרוקנת U1.
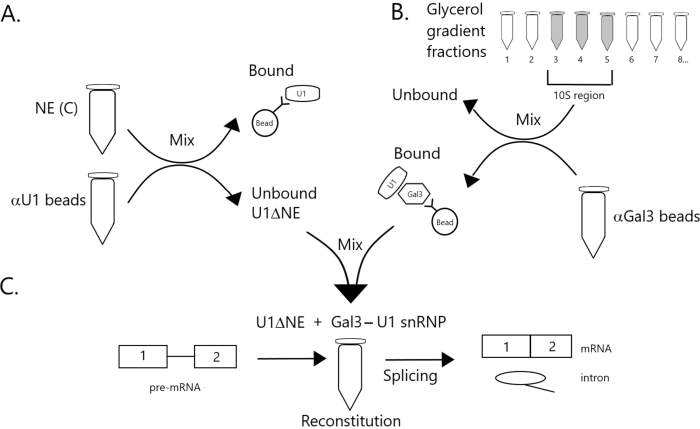
איור 1: תרשים סכמטי הממחיש את ההשלמה של פעילות ההצמדה בתמצית גרעינית מרוקנת מ- U1 snRNP על ידי קומפלקס Gal3-U1 snRNP על חרוזים. (A) NE במאגר C (NE(C)) הוא דגירה עם חרוזי חלבון A-Sepharose בשילוב עם נוגדי U1 snRNP (חרוזים αU1). השבר הלא מאוגד מתרוקן מ- U1 snRNP (U1ΔNE). (B) NE במאגר D (NE(D)) מחולק מעבר הדרגתי של 12%-32% גליצריל על-ידי אולטרה-צנטריפוגה. שברים המתאימים לאזור 10S (שברים 3-5) משולבים ומעורבבים עם חרוזים בשילוב עם נוגדנים נגד Gal3 (חרוזים αGal3). החומר הקשור החרוזים מכיל מונופרקל SnRNP 3-U1. (ג) מתחם Gal3-U1 snRNP מחלק (B) מעורבב עם U1ΔNE מחלק (A) בבדיקה משולבת באמצעות מצע קדם-mRNA של MINX עם תווית 32P והמתווכים והמוצרים של תגובת ההצמדה מנותחים על ידי אלקטרופורזה ג’ל ואוטרדיוגרפיה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
