إنتاج معظم RNAS رسول eukaryotic (mRNAs) ينطوي على إزالة introns وربط exons في عملية نووية يطلق عليها اسم ما قبل مرنا الربط1. نوعين من مجمعات بروتين الحمض النووي الريبي (RNPs) توجيه معالجة الحمض النووي الريبي قبل رسول في ميرنا ناضجة عبر مجمعات الطحال. يتم تشكيل فئة واحدة ، RNPs ما قبل الرسول الوليدة ، نسخا مشتركة من خلال ربط بروتينات RNP النووية غير المتجانسة وغيرها من البروتينات الملزمة للجيش الملكي النيبالي ، بما في ذلك بعض أفراد عائلة SR ، مما ينتج مجمعات hnRNP2. ويرتبط من الدرجة الثانية، RNPs النووية الصغيرة الغنية uracil (U snRNPs مع U1، U2، U4، U5، وU6 snRNAs) مع U محددة والبروتينات الأساسية3،4. تتفاعل UnRNPs بطريقة مرتبة مع مناطق محددة من RNPs ما قبل الرسول في مسار إعادة عرض ديناميكي حيث يتم استئصال السترونز وربط exons لإنتاج mRNPs5 ناضجة. ويشارك العديد من البروتينات النووية الإضافية في أحداث المعالجة هذه6.
Galectin-1 (Gal1) و galectin-3 (Gal3) هما بروتينان مطلوبان في مسار الربط كما هو موضح في دراسات إعادة تشكيل الاستنفاد7،8. إزالة كل من galectins من الربط المستخلصات النووية المختصة (NE) يلغي التجميع الربط ونشاط الربط في خطوة مبكرة. إضافة أي galectin إلى مثل هذه NE المنضب بشكل مضاعف يستعيد كلا النشاطين. Gal1 و Gal3 هي مكونات اللصقات النشطة كما يتضح من السبق المناعي المحدد لمرنا ما قبل، وسيطة الربط، ورنا ناضجة من قبل antiserum محددة إما Gal1 أو Gal39. الأهم من ذلك، Gal3 المنتسبين مع SnRNA U الذاتية التي تحتوي على جزيئات في NE خارج مسار الربط كما هو مبين من خلال هطول الأمطار من snRNPs من قبل antisera10 المضادة Gal3. وأخيرا، إسكات Gal3 في خلايا هيلا يغير أنماط الربط من الجينات العديدة11.
في NE قبل احتضان لتفكيك اللصقات مسبقة الشكل12، تم العثور على snRNPs في مجمعات متعددة الرسوب في تدرجات الجلسرين من 7S إلى أكبر من 60S. على الرغم من أن الكسر التدرج الجلسرين هو تقنية شائعة لعزل المجمعات والمكونات الطحال (انظر المراجع13,14,15 على سبيل المثال), لقد قمنا بتوسيع هذه الطريقة عن طريق توصيف كسور محددة باستخدام الأجسام المضادة المناعية. وsnRNP الرسوب في 10S يحتوي فقط U1 snRNA جنبا إلى جنب مع Gal3. إن التكسير المناعي للكسر 10S مع antisera محدد ل Gal3 أو U1 snRNP يشارك في التعجيل بكل من U1 و Gal3 مما يشير إلى أن بعض الجسيمات الأحادية U1 snRNP لا بد أن تكون Gal310. كما U1 snRNP هو أول مجمع يربط إلى ما قبل mRNP في التجميع spliceosomal1،5، تمثل هذه الخطوة موقع دخول محتمل لGal3 في مسار الربط. على هذا الأساس، أظهرنا أن 10S Gal3-U1 snRNP أحادية الجسيمات ملزمة المضادة Gal3 التي تحتوي على الخرز استعادة نشاط الربط إلى U1 snRNP استنفدت NE، وإنشاء هذا المجمع كآلية واحدة التي يتم تجنيد Gal3 في المسار الربط16. وهذا يتناقض مع محاولات عزل اللصقات في مراحل محددة في رد فعل الربط وفهرسة العوامل المرتبطة بها17,18. وفي مثل هذه الدراسات، يتم التأكد من وجود عوامل معينة في وقت ما ولكن ليس الآلية التي تم تحميلها بها.
كنا قد وصفنا بالتفصيل في إعداد NE، وركيزة الربط، وتجميع خليط رد فعل الربط، وتحليل المنتجات في توثيقنا لدور galectins في الربط قبل ميرنا19. ونصف الآن الإجراءات التجريبية لتجزئ المستخلصات النووية للحصول على جزء مثري في مجمع Gal3 – U1 snRNP ولانتقاء المناعة للمجمع الأخير لإعادة تشكيل نشاط الربط في مستخلص نووي مستنفد U1.
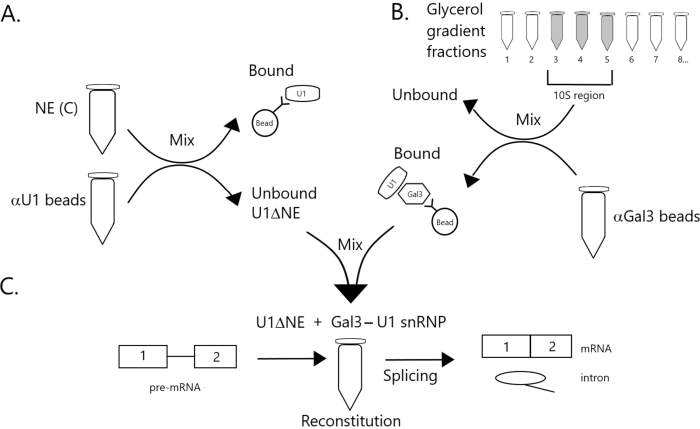
الشكل 1: مخطط تخطيطي يوضح استكمال نشاط الربط في استخراج نووي مستنزف من U1 snRNP بواسطة مجمع Gal3-U1 snRNP على الخرز. (A) NE في العازل C (NE(C)) يتم احتضانه بخرز البروتين A-Sepharose مقترنا بشكل متناقض مع حبات SnRNP المضادة U1 (حبات αU1). يتم استنفاد الكسر غير منضم من U1 snRNP (U1ΔNE). (ب) يتم تجزئة NE في المخزن المؤقت D (NE(D)) على تدرج جلسيرول 12٪-32٪ عن طريق الطرد المركزي الفائق. يتم الجمع بين الكسور المقابلة لمنطقة 10S (الكسور 3-5) وخلطها مع الخرز المقترن بشكل مشترك مع الأجسام المضادة المضادة ل Gal3 (حبات αGal3). المواد ملزمة الخرز يحتوي على Gal3-U1 snRNP أحادية الحزبية. (ج) يتم خلط مجمع Gal3-U1 snRNP من الجزء (B) مع U1ΔNE من الجزء (A) في مقايسة الربط باستخدام ركيزة MINX قبل ميرنا المسماة 32P ويتم تحليل وسيطة ومنتجات تفاعل الربط بواسطة الكتروفلوريس الهلامي والتصوير الإشعاعي التلقائي. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
