Produksjon av de fleste eukaryote budbringere RNAer (mRNAer) innebærer fjerning av introner og ligasjon av eksoner i en kjernefysisk prosess kalt pre-mRNA skjøting1. To klasser av RNA-proteinkomplekser (RNPer) styrer behandlingen av pre-messenger RNA til moden mRNA via spleiseosomale komplekser. En klasse, nascent pre-messenger RNPs, dannes co-transkripsjonelt ved binding av heterogene kjernefysiske RNP-proteiner og andre RNA-bindende proteiner, inkludert noen medlemmer av SR-familien, noe som gir hnRNP-komplekser2. Den andre klassen, uracil-rike små kjernefysiske RNPer (U snRNPer med U1, U2, U4, U5 og U6 snRNAs) er forbundet med U-spesifikke og kjerneproteiner3,4. De amerikanske snRNPene samhandler på en ordnet måte med spesifikke regioner av pre-messenger RNPer i en dynamisk ombyggingsvei som introner utskilles og exons er ligated å produsere modne mRNPs5. Mange andre kjerneproteiner deltar i disse behandlingshendelsene6.
Galectin-1 (Gal1) og galectin-3 (Gal3) er to proteiner som er nødvendige faktorer i skjøteveien som vist ved uttømming-rekonstitueringsstudier7,8. Fjerning av begge galectins fra skjøting kompetente kjernefysiske ekstrakter (NE) avskaffer skjøteosom montering og skjøting aktivitet på et tidlig trinn. Tillegg av begge galectin til en slik dobbelt utarmet NE gjenoppretter begge aktivitetene. Gal1 og Gal3 er komponenter av aktive skjøteosomer som det fremgår av spesifikk immunoprecipitation av pre-mRNA, skjøting mellomprodukter, og moden mRNA ved antiserum spesifikk for enten Gal1 eller Gal39. Viktigst, Gal3 forbinder med endogene U snRNA inneholder partikler i NE utenfor skjøteveien som vist ved nedbør av snRNPs av anti-Gal3 antisera10. Til slutt endrer silencing av Gal3 i HeLa-celler skjøtemønstre av mange gener11.
I NE pre-inkubert for å demontere preformede skjøteosomer12, snRNPs finnes i flere komplekser sedimentering i glyserol gradienter fra 7S til større enn 60S. Selv om glyserol gradient fraksjonering er en vanlig teknikk for isolering av spleiseosomale komplekser og komponenter (se referanser13,14,15 for eksempel), har vi utvidet denne metoden ved å karakterisere spesifikke fraksjoner ved hjelp av antistoff immunoprecipitations. En snRNP sedimentering ved 10S inneholder bare U1 snRNA sammen med Gal3. Immunoprecipitation av 10S fraksjonen med antisera spesifikk for Gal3 eller U1 snRNP co-utfelling både U1 og Gal3 indikerer noen av U1 snRNP monopartikler er bundet til Gal310. Siden U1 snRNP er det første komplekset som binder seg til pre-mRNP i spleiseosomal montering1,5, representerer dette trinnet et potensielt inngangssted for Gal3 inn i skjøtebanen. På dette grunnlaget viste vi at 10S Gal3-U1 snRNP monopartikler bundet til anti-Gal3 som inneholder perler restaurert skjøteaktivitet til en U1 snRNP utarmet NE, og etablerer dette komplekset som en mekanisme som Gal3 rekrutteres til den spleiseosomale banen16. Dette står i kontrast til forsøk på å isolere skjøteosomer på bestemte stadier i skjøtereaksjonen og katalogisere de tilhørende faktorene17,18. I slike studier er tilstedeværelsen av visse faktorer på et tidspunkt fastslått, men ikke mekanismen de ble lastet på.
Vi hadde tidligere beskrevet i detalj utarbeidelsen av NE, skjøteunderlaget, monteringen av skjøtereaksjonsblandingen og analysen av produkter i vår dokumentasjon av galektinenes rolle i pre-mRNA skjøting19. Vi beskriver nå de eksperimentelle prosedyrene for fraksjonering av atomekstrakter for å oppnå en brøkdel beriket i Gal3 – U1 snRNP-kompleks og for immunvalg av sistnevnte kompleks for å rekonstituere skjøteaktivitet i et U1-utarmet atomekstrakt.
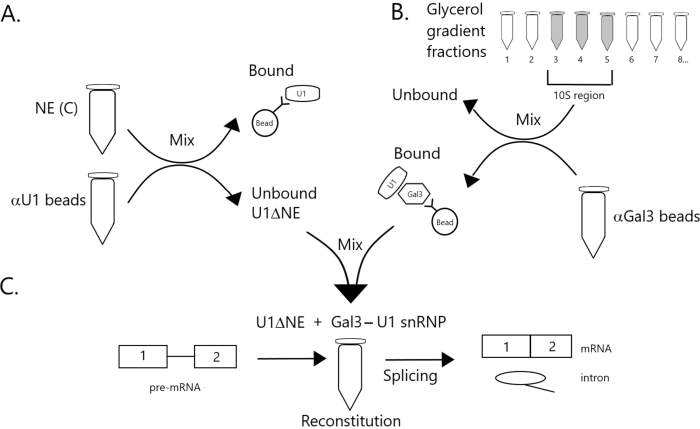
Figur 1: Skjematisk diagram som illustrerer komplementering av skjøteaktivitet i kjernefysisk ekstrakt utarmet av U1 snRNP av et Gal3-U1 snRNP-kompleks på perler. (A) NE i Buffer C (NE(C)) inkuberes med Protein A-Sepharose perler kovalent kombinert med anti-U1 snRNP (αU1 perler). Den ubundne brøkdelen er utarmet av U1 snRNP (U1ΔNE). (B) NE i buffer D (NE(D)) er fraksjonert over en 12%-32% glyserol gradient ved ultracentrifugation. Fraksjoner som tilsvarer 10S-regionen (brøker 3-5) kombineres og blandes med perler kovalent kombinert med anti-Gal3 antistoffer (αGal3 perler). Materialet bundet til perlene inneholder en Gal3-U1 snRNP monopartikkel. (C) Gal3-U1 snRNP-komplekset fra part (B) blandes med U1ΔNE fra del (A) i en skjøteanalyse ved hjelp av 32P-merket MINX pre-mRNA-substrat og mellomprodukter og produkter av skjøtereaksjonen analyseres ved gelelektroforese og autoradiografi. Klikk her for å se en større versjon av denne figuren.
