A produção da maioria dos RNAs (mRNAs) do mensageiro eucariótico envolve a remoção de introns e a ligadura de exons em um processo nuclear chamado de emenda pré-mRNA1. Duas classes de complexos de proteína RNA (RNPs) direcionam o processamento do RNA pré-mensageiro em mRNA maduro através de complexos spliceossômicos. Uma classe, rnps pré-mensageiro nascente, é formada co-transcrição pela vinculação de proteínas heterogêneas nucleares RNP e outras proteínas de ligação de RNA, incluindo alguns membros da família SR, produzindo complexos hnRNP2. A segunda classe, rica em uracil, pequenas RNPs nucleares (U snRNPs com U1, U2, U4, U5 e U6 snRNAs) está associada com proteínas específicas e core u3,4. Os SNRNPs da U interagem de forma ordenada com regiões específicas de RNPs pré-mensageiros em um caminho de remodelação dinâmica à medida que os introns são extirpados e exons são ligados para produzir mRNPs5 maduros. Muitas proteínas nucleares adicionais participam desses eventos de processamento6.
Galectin-1 (Gal1) e galectin-3 (Gal3) são duas proteínas que são fatores necessários na via de emenda, como mostrado pelos estudos de esgotamento-reconstituição7,8. A remoção de ambas as galectinas de emendas extratos nucleares competentes (NE) abolia a montagem de emendas e a atividade de emenda em um passo inicial. A adição de uma galectina a um NE duplamente esgotado restaura ambas as atividades. Gal1 e Gal3 são componentes de emendas ativas, evidenciadas por imunoprecipitação específica de pré-mRNA, intermediários de emenda e mRNA maduro por antiserum específico para Gal1 ou Gal39. É importante ressaltar que a Gal3 associa-se ao snRNA u endógeno contendo partículas no NE fora da via de emenda, como mostrado pela precipitação de snRNPs por anti-Gal3 antisera10. Finalmente, o silenciamento de Gal3 em células HeLa altera padrões de emenda de numerosos genes11.
No NE pré-incubado para desmontar os spliceosomes pré-formados12, snRNPs são encontrados em múltiplos complexos sedimentando em gradientes de glicerol de 7S para maiores de 60S. Embora o fracionamento do gradiente de glicerol seja uma técnica comum para o isolamento de complexos e componentes spliceossômicos (ver referências13,14,15 por exemplo), ampliamos esse método caracterizando frações específicas usando imunoprecipitações de anticorpos. Um sedimento snRNP em 10S contém apenas u1 snRNA juntamente com Gal3. A imunoprecipitação da fração 10S com antisera específica para Gal3 ou U1 snRNP co-precipita tanto u1 quanto Gal3 indicando que algumas das mon partículas do U1 snRNP estão ligadas a Gal310. Como o U1 snRNP é o primeiro complexo que se liga ao pré-mRNP em conjuntos emendasomais1,5, esta etapa representa um potencial local de entrada para Gal3 na via de emenda. Com base nisso, mostramos que as monpartículas 10S Gal3-U1 snRNP ligadas à anti-Gal3 contendo contas restauradas para um NE esgotado U1 snRNP, estabelecendo este complexo como um mecanismo pelo qual Gal3 é recrutado para o caminho emendalomal16. Isso contrasta com as tentativas de isolar os emendas em estágios específicos na reação de emenda e catalogação dos fatores associados17,18. Em tais estudos, a presença de certos fatores em algum momento é apurada, mas não o mecanismo pelo qual foram carregados.
Havia descrito anteriormente em detalhes a preparação do NE, o substrato de emenda, a montagem da mistura de reação de emenda, e a análise de produtos em nossa documentação do papel das galectinas no splicing pré-mRNA19. Descrevemos agora os procedimentos experimentais para o fracionamento de extratos nucleares para obter uma fração enriquecida no complexo Gal3 – U1 snRNP e para a imuno-seleção deste último complexo para reconstituir a atividade de emenda em um extrato nuclear esgotado pelo U1.
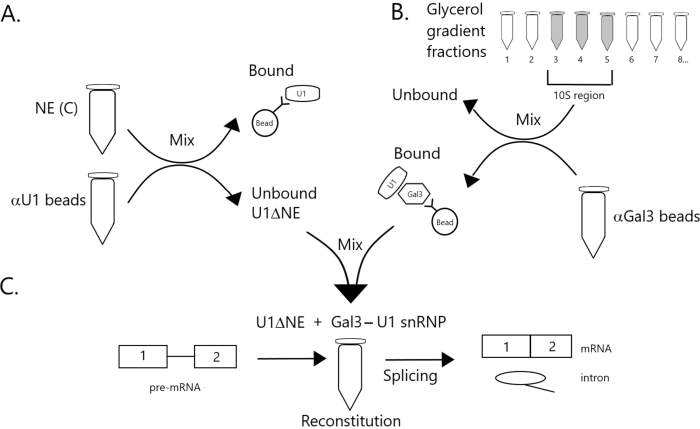
Figura 1: Diagrama esquemático ilustrando a complementação da atividade de emenda em extrato nuclear esgotado de U1 snRNP por um complexo de snRNP Gal3-U1 em contas. (A) NE em Buffer C (NE(C)) é incubado com contas de proteína A-Sepharose covalentemente acopladas com anti-U1 snRNP (contas αU1). A fração desvinculada está esgotada de U1 snRNP (U1ΔNE). (B) NE em Buffer D (NE(D)) é fracionado sobre um gradiente de glicerol de 12%-32% por ultracentrifugação. Frações correspondentes à região 10S (frações 3-5) são combinadas e misturadas com contas covalentemente acopladas com anticorpos anti-Gal3 (contas αGal3). O material ligado às contas contém uma monopartícula de SnRNP Gal3-U1. (C) O complexo de snRNP Gal3-U1 da Parte (B) é misturado com U1ΔNE da Parte (A) em um ensaio de emenda usando substrato minx pré-mRNA com 32P e os intermediários e produtos da reação de emenda são analisados por eletroforese gel e autoradiografia. Clique aqui para ver uma versão maior desta figura.
