Produktion af de fleste eukaryote messenger RNA’er (mRNAs) indebærer fjernelse af introner og ligation af exons i en nuklear proces, der kaldes præ-mRNA splejsning1. To klasser af RNA-proteinkomplekser (RRNPs) leder behandlingen af RNA før budbringeren ind i moden mRNA via splejsede komplekser. En klasse, spirende pre-messenger RNPs, er dannet co-transcriptionally ved binding af heterogene nukleare RNP proteiner og andre RNA-bindende proteiner, herunder nogle medlemmer af SR familien, der giver hnRNP komplekser2. Den anden klasse, uracil-rige små nukleare RNPs (U snRNPs med U1, U2, U4, U5, og U6 snRNAs) er forbundet med U-specifikke og kerneproteiner3,4. U snRNPs interagerer på en ordnet måde med specifikke regioner af præ-messenger RNPs i en dynamisk remodeling vej som introns er fjernet og exons er ligated til at producere modne mRNPs5. Mange yderligere nukleare proteiner deltager i disse behandlingshændelser6.
Galectin-1 (Gal1) og galectin-3 (Gal3) er to proteiner, der er nødvendige faktorer i splejsning vej som vist ved udtømning-rekonstitution undersøgelser7,8. Fjernelse af begge galectiner fra splejsning af kompetente nukleare ekstrakter (NE) afskaffer splejsning og splejsning aktivitet på et tidligt trin. Tilføjelse af enten galectin til en sådan dobbelt udtømt NE genopretter begge aktiviteter. Gal1 og Gal3 er komponenter i aktive splekviomer, som det fremgår af specifik immunprecipitation af præ-mRNA, splejsning mellemprodukter, og modne mRNA ved antiserum specifikke for enten Gal1 eller Gal39. Det er vigtigt, gal3 forbinder med endogene U snRNA indeholdende partikler i NE uden for splejsning vej som det fremgår af nedbør af snRNPs af anti-Gal3 antisera10. Endelig ændrer lyddæmpning af Gal3 i HeLa-celler splejsningsmønstre af mange gener11.
I NE præ-inkuberet til at adskille præformede spleknoomer12, snRNPs findes i flere komplekser sedimentering i glycerol gradienter fra 7S til større end 60S. Selvom glycerolgradientfraktionering er en almindelig teknik til isolering af splejsede komplekser og komponenter (se f.eks. referencer13,14,15), har vi udvidet denne metode ved at karakterisere specifikke fraktioner ved hjælp af antistofimmunpitationer. En snRNP sedimentering ved 10S indeholder kun U1 snRNA sammen med Gal3. Immunprecipitation af 10S-fraktionen med antisera, der er specifik for Gal3 eller U1 snRNP, medudfælder både U1 og Gal3, hvilket indikerer, at nogle af U1 snRNP monopartiklerne er bundet til Gal310. Da U1 snRNP er det første kompleks, der binder sig til præ-mRNP i splejset samling1,5, repræsenterer dette trin et potentielt indgangssted for Gal3 i splejsningsvejen. På dette grundlag viste vi, at 10S Gal3-U1 snRNP monopartikler bundet til anti-Gal3 indeholder perler restaureret splejsning aktivitet til en U1 snRNP udtømt NE, oprettelse af dette kompleks som en mekanisme, hvorved Gal3 er rekrutteret i splejset vej16. Dette står i kontrast til forsøg på at isolere splejsomer på bestemte stadier i splejsning reaktion og katalogisering de tilhørende faktorer17,18. I sådanne undersøgelser fastslås tilstedeværelsen af visse faktorer på et tidspunkt, men ikke den mekanisme, hvormed de blev indlæst.
Vi havde tidligere beskrevet i detaljer forberedelsen af NE, splejsningsubstratet, samlingen af splejsning reaktion blanding, og analysen af produkter i vores dokumentation af galectins rolle i præ-mRNA splejsning19. Vi beskriver nu forsøgsprocedurerne for fraktionering af nukleare ekstrakter for at opnå en brøkdel beriget i Gal3 – U1 snRNP-kompleks og for immunvalg af sidstnævnte kompleks for at rekonstruere splejsningsaktivitet i et U1-udtømt nukleart ekstrakt.
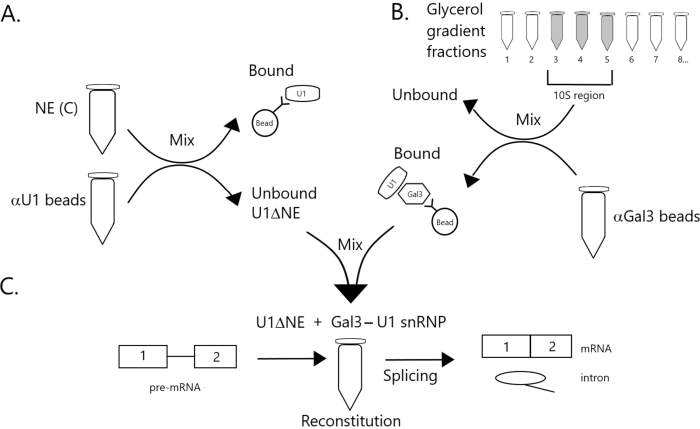
Figur 1: Skemat diagram, der illustrerer komplementering af splejsning i nukleart ekstrakt udtømt af U1 snRNP af et Gal3-U1 snRNP-kompleks på perler. (A) NE i Buffer C (NE(C)) inkuberes med Protein A-Sepharose perler kovalent kombineret med anti-U1 snRNP (αU1 perler). Den ubundne brøk er udtømt af U1 snRNP (U1ΔNE). (B) NE i Buffer D (NE(D)) er fraktioneret over en 12%-32% glycerol gradient ved ultracentrifugering. Fraktioner svarende til 10S-regionen (fraktioner 3-5) kombineres og blandes med perler kovalent kombineret med anti-Gal3 antistoffer (αGal3 perler). Materialet bundet til perlerne indeholder en Gal3-U1 snRNP monopartikler. (C) Gal3-U1 snRNP-komplekset fra del (B) blandes med U1ΔNE fra del (A) i en splejsning ved hjælp af 32P-mærket MINX pre-mRNA-substrat, og mellemprodukter og produkter fra splejsningreaktionen analyseres ved gelelektroforse og autoradiografi. Klik her for at se en større version af dette tal.
