Immuuncellen verdedigen zich tegen ziekteverwekkers en kankercellen door voortdurend de oppervlakken van doelcellen te kruipen en te scannen op antigenen, waarbij hun oppervlak 1,2 wordt gestudeerd. Antigeenherkenning wordt geïnitieerd na binding tussen de T-celreceptor (TCR) en het peptide-major histocompatibiliteitscomplex MHC (pMHC) -complex dat tot expressie komt op het oppervlak van doelcellen. Omdat TCR-pMHC-herkenning plaatsvindt op de kruising tussen twee mobiele cellen, wordt al lang vermoed dat het mechanische krachten ervaart. Bovendien leidde dit tot het mechanosensormodel van TCR-activering, dat suggereert dat TCR-krachten bijdragen aan zijn functie 3,4. Om te begrijpen wanneer, waar en hoe mechanische krachten bijdragen aan de T-celfunctie, is het noodzakelijk om hulpmiddelen te ontwikkelen om de moleculaire krachten die door T-cellen worden overgedragen te visualiseren. Traditioneel worden methoden zoals tractiekrachtmicroscopie (TFM) en micropillar arrays gebruikt om cellulaire krachten 5,6 te onderzoeken. De krachtgevoeligheid van TFM en micropilaire arrays bevindt zich echter op de nanonewton (nN) schaal en is dus vaak onvoldoende om de moleculaire piconewton (pN) krachten te bestuderen die door celreceptorenworden overgedragen 7. Om de kracht en ruimtelijke resolutie voor detectie te verbeteren, pionierde ons laboratorium met de ontwikkeling van moleculaire spanningsondes, die aanvankelijk werden gesynthetiseerd met behulp van polyethyleenglycol (PEG) polymeren7. Moleculaire spanningsondes bestaan uit een uitschuifbare moleculaire “veer” (PEG, eiwit, DNA) geflankeerd door een fluorofoor en quencher en zijn verankerd op een oppervlak. Krachten die op het eindpunt van de sonde worden uitgeoefend, leiden tot de uitbreiding ervan, waardoor de fluorofoor en de quencher worden gescheiden en zo een sterk fluorescentiesignaal wordt gegenereerd (figuur 1A)8,9,10.
In het afgelopen decennium hebben we een bibliotheek ontwikkeld van verschillende klassen van moleculaire spanningsondes met veerelementen gemaakt van nucleïnezuren11, eiwitten10 en polymeren8. Onder deze bieden de op DNA gebaseerde spanningssondes de hoogste signaal-ruisverhouding en de grootste krachtgevoeligheid, die gemakkelijk kan worden afgestemd van een paar pN tot ~ 20 pN11. We hebben deze real-time DNA-spanningsondes gebruikt om de moleculaire krachten te bestuderen die worden gegenereerd door vele verschillende celtypen, waaronder fibroblasten, kankercellen, bloedplaatjes en immuuncellen11,12,13. Dit manuscript zal protocollen beschrijven voor het synthetiseren en assembleren van DNA-spanningsondes op een oppervlak om moleculaire receptorkrachten in kaart te brengen met pN-krachtresolutie met behulp van een conventionele fluorescentiemicroscoop. Hoewel de huidige procedure chemische modificaties aan het nucleïnezuur omvat om de fluorescerende reporter te introduceren (figuur 1B), is het belangrijk op te merken dat veel van de modificatie- en zuiveringsstappen kunnen worden uitbesteed aan aangepaste DNA-synthesebedrijven. Daarom is de technologie van DNA-spanningsondes gemakkelijk en toegankelijk voor de bredere celbiologie en mechanobiologiegemeenschappen.
Kortom, om DNA-spanningssensoren samen te stellen, wordt een DNA-haarspeld gehybridiseerd tot een fluorescerende ligandstreng op de ene arm en een quencher-ankerstreng op de andere arm en vervolgens geïmmobiliseerd op een glazen substraat (figuur 1C, real-time spanning). Bij afwezigheid van mechanische kracht wordt de haarspeld gesloten en wordt dus de fluorescentie geblust. Wanneer de uitgeoefende mechanische kracht echter groter is dan de F1/2 (de kracht bij evenwicht die leidt tot een kans van 50% om zich te ontvouwen), smelt de haarspeld mechanisch en wordt een fluorescerend signaal gegenereerd.
Voortbouwend op de real-time DNA-spanningssensor beschrijven we ook protocollen om geaccumuleerde krachten in kaart te brengen, wat vooral nuttig is voor het bestuderen van interacties tussen receptoren op immuuncellen en hun natuurlijke ligand. Dit komt omdat immuunreceptoren vaak kortstondige bindingen vertonen 3,14. Geaccumuleerde krachten worden in beeld gebracht met behulp van een “vergrendelingsstreng” die bij voorkeur bindt aan open DNA-haarspelden en de opslag van fluorescentiesignalen mogelijk maakt die verband houden met mechanische trekgebeurtenissen (figuur 1C, vergrendelde spanning). De vergrendelingsstreng is ontworpen om een cryptische bindingsplaats te binden die wordt blootgesteld aan mechanisch geïnduceerd smelten van de haarspeld en de haarspeld in de open toestand te vergrendelen door het opnieuw vouwen van de haarspeld te blokkeren, waardoor het spanningssignaal wordt opgeslagen en een geaccumuleerde spanningskaart wordt gegenereerd. Bovendien is de vergrendelingsstreng ontworpen met een acht-nucleotide teengreep, die een teengreep-gemedieerde strengverplaatsingsreactie mogelijk maakt met zijn volledige complement, de “ontgrendelende” streng. Met de toevoeging van de ontgrendelingsstreng wordt de gebonden vergrendelingsstreng van de haarspeldconstructie gestript, waardoor het opgeslagen spanningssignaal wordt gewist en de haarspeld wordt teruggezet naar de real-time status.
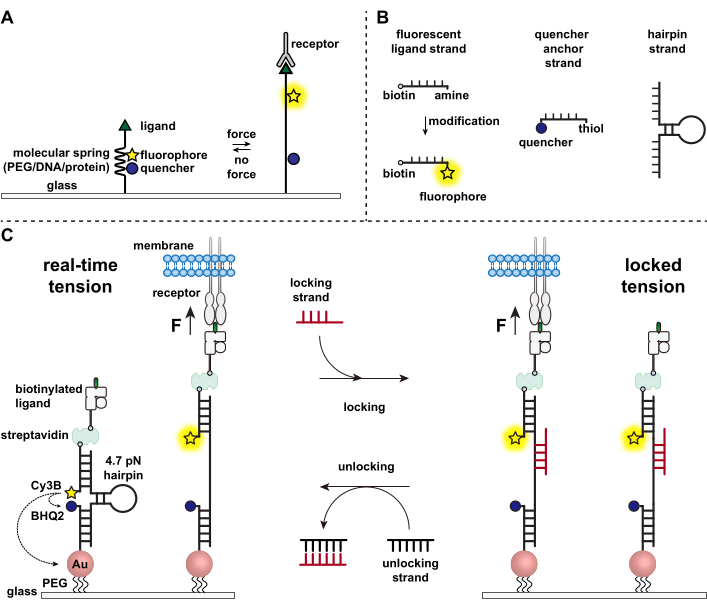
Figuur 1: Schema van de state-of-art moleculaire spanningsondes . (A) Algemeen ontwerp van real-time moleculaire spanningsonde, (B) Strengen voor de DNA-gebaseerde spanningsondeconstructie, en (C) gemanipuleerde DNA-gebaseerde spanningsondes en hun schakelen tussen real-time toestand en vergrendelde toestand. Klik hier om een grotere versie van deze figuur te bekijken.
Het hoofdprotocol bestaat uit vier hoofdsecties – oligonucleotidepreparaat, oppervlaktevoorbereiding, beeldvorming en gegevensanalyse. Dit protocol is met succes aangetoond door ons laboratorium en anderen in naïeve en geactiveerde OT-1 CD8 + T-cellen, OT-II CD4 + -cellen, evenals hybridomen, en kan worden toegepast om verschillende immuuncelreceptoren te ondervragen, waaronder T-celreceptor, geprogrammeerde celdoodreceptor (PD1) en lymfocytfunctie-geassocieerde antigeen 1 (LFA-1) krachten. OT-1 CD8+ naïeve T-cellen worden in dit artikel als voorbeeldcellijn gebruikt.
