Bağışıklık hücreleri, hedef hücrelerin yüzeylerini antijenler için sürekli tarayarak ve tarayarak, yüzeylerini 1,2 saplayarak patojenlere ve kanser hücrelerine karşı savunurlar. Antijen tanıma, hedef hücrelerin yüzeyinde eksprese edilen T hücre reseptörü (TCR) ile peptid-majör histokompatibilite kompleksi MHC (pMHC) kompleksi arasındaki bağlanma ile başlatılır. TCR-pMHC tanıma iki mobil hücre arasındaki kavşakta meydana geldiğinden, uzun zamandır mekanik kuvvetler yaşadığından şüphelenilmektedir. Dahası, bu, TCR kuvvetlerinin 3,4 işlevine katkıda bulunduğunu öne süren TCR aktivasyonunun mekanosensör modeline yol açtı. Mekanik kuvvetlerin T hücresi fonksiyonuna ne zaman, nerede ve nasıl katkıda bulunduğunu anlamak için, T hücreleri tarafından iletilen moleküler kuvvetleri görselleştirmek için araçlar geliştirmek zorunludur. Geleneksel olarak, çekiş kuvveti mikroskobu (TFM) ve mikrosütun dizileri gibi yöntemler, hücresel kuvvetleri araştırmak için kullanılır 5,6. Bununla birlikte, TFM ve mikrosütun dizilerinin kuvvet duyarlılığı nanonewton (nN) ölçeğindedir ve bu nedenle hücre reseptörleri tarafından iletilen moleküler pikonewton (pN) kuvvetlerini incelemek için genellikle yetersizdir7. Tespit için kuvveti ve uzamsal çözünürlüğü iyileştirmek için laboratuvarımız başlangıçta polietilen glikol (PEG) polimerleri7 kullanılarak sentezlenen moleküler gerilim problarının geliştirilmesine öncülük etti. Moleküler gerilim probları, bir florofor ve söndürücü ile çevrili uzatılabilir bir moleküler “yay” dan (PEG, protein, DNA) oluşur ve bir yüzeye sabitlenir. Probun ucuna uygulanan kuvvetler, florofor ve söndürücüyü ayırarak genişlemesine yol açar ve böylece güçlü bir floresan sinyali üretir (Şekil 1A)8,9,10.
Son on yılda, nükleik asitler11, proteinler10 vepolimerler 8’den yapılmış yay elementleri ile farklı moleküler gerilim probları sınıflarından oluşan bir kütüphane geliştirdik. Bunlar arasında, DNA tabanlı gerilim probları, birkaç pN’den ~ 20 pN11’e kadar kolayca ayarlanabilen en yüksek sinyal-gürültü oranını ve en yüksek kuvvet hassasiyetini sağlar. Bu gerçek zamanlı DNA gerilim problarını, fibroblastlar, kanser hücreleri, trombositler ve bağışıklık hücreleri11,12,13 dahil olmak üzere birçok farklı hücre tipi tarafından üretilen moleküler kuvvetleri incelemek için kullandık. Bu makalede, geleneksel floresan mikroskobu kullanılarak moleküler reseptör kuvvetlerini pN kuvvet çözünürlüğü ile haritalamak için DNA gerilim problarını bir yüzey üzerinde sentezlemek ve birleştirmek için kullanılan protokoller anlatılacaktır. Mevcut prosedür, floresan muhabiri tanıtmak için nükleik asitte kimyasal modifikasyonlar içerirken (Şekil 1B), modifikasyon ve saflaştırma adımlarının çoğunun özel DNA sentez şirketlerine dış kaynaklı olabileceğini belirtmek önemlidir. Bu nedenle, DNA gerilim probları teknolojisi kolaydır ve daha geniş hücre biyolojisi ve mekanobiyoloji toplulukları için erişilebilirdir.
Kısaca, DNA gerilim sensörlerini monte etmek için, bir DNA saç tokası bir koldaki floresan ligand ipliğine ve diğer koldaki bir söndürücü çapa ipliğine hibridize edilir ve daha sonra bir cam substrat üzerinde hareketsiz hale getirilir (Şekil 1C, gerçek zamanlı gerginlik). Mekanik kuvvetin yokluğunda, saç tokası kapatılır ve böylece floresan söndürülür. Bununla birlikte, uygulanan mekanik kuvvet F1/2’den (% 50’lik bir açılma olasılığına yol açan dengedeki kuvvet) daha büyük olduğunda, saç tokası mekanik olarak erir ve bir floresan sinyali üretilir.
Gerçek zamanlı DNA gerilim sensörüne dayanarak, birikmiş kuvvetleri haritalamak için protokolleri de açıklıyoruz; bu, bağışıklık hücreleri üzerindeki reseptörler ile doğal ligandları arasındaki etkileşimleri incelemek için özellikle yararlıdır. Bunun nedeni, bağışıklık reseptörlerinin sıklıkla kısa ömürlü bağlar göstermesidir 3,14. Biriken kuvvetler, tercihen açık DNA saç tokalarına bağlanan ve mekanik çekme olaylarıyla ilişkili floresan sinyallerinin depolanmasına izin veren bir “kilitleme” ipliği kullanılarak görüntülenir (Şekil 1C, kilitli gerilim). Kilitleme ipliği, saç tokasının mekanik olarak indüklenen erimesi üzerine açığa çıkan şifreli bir bağlama bölgesini bağlamak ve saç tokasının yeniden katlanmasını engelleyerek saç tokasını açık durumda kilitlemek, böylece gerginlik sinyalini depolamak ve birikmiş bir gerilim haritası oluşturmak için tasarlanmıştır. Dahası, kilitleme ipliği, tam tamamlayıcısı olan “kilit açma” ipliği ile ayak parmağı aracılı bir iplik yer değiştirme reaksiyonu sağlayan sekiz nükleotid ayak parmağı ile tasarlanmıştır. Kilit açma telinin eklenmesiyle, bağlı kilitleme teli saç tokası yapısından sıyrılır, depolanan gerginlik sinyali silinir ve saç tokasını gerçek zamanlı duruma geri döndürür.
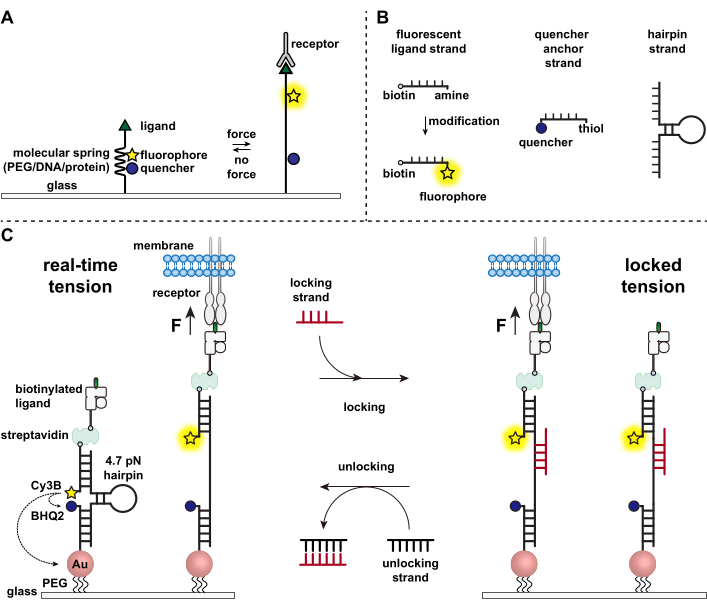
Resim 1: Son teknoloji moleküler gerilim problarının şeması . (A) Gerçek zamanlı moleküler gerilim probunun genel tasarımı, (B) DNA tabanlı gerilim probu yapısı için iplikçikler ve (C) DNA tabanlı gerilim probları ve bunların gerçek zamanlı durum ile kilitli durum arasında geçiş yapması. Bu şeklin daha büyük bir versiyonunu görmek için lütfen buraya tıklayın.
Ana protokol dört ana bölümden oluşur – oligonükleotid hazırlama, yüzey hazırlama, görüntüleme ve veri analizi. Bu protokol, laboratuvarımız ve diğerleri tarafından naif ve aktive edilmiş OT-1 CD8 + T hücreleri, OT-II CD4 + hücreleri ve hibridomlarda başarıyla gösterilmiştir ve T hücre reseptörü, programlanmış hücre ölüm reseptörü (PD1) ve lenfosit fonksiyonu ile ilişkili antijen 1 (LFA-1) kuvvetleri dahil olmak üzere farklı bağışıklık hücresi reseptörlerini sorgulamak için uygulanabilir. OT-1 CD8+ naif T hücreleri bu yazıda örnek hücre çizgisi olarak kullanılmıştır.
