Le cellule immunitarie difendono dagli agenti patogeni e dalle cellule tumorali strisciando e scansionando continuamente le superfici delle cellule bersaglio alla ricerca di antigeni, costellando la loro superficie 1,2. Il riconoscimento dell’antigene viene avviato al momento del legame tra il recettore delle cellule T (TCR) e il complesso di istocompatibilità peptide-maggiore MHC (pMHC) espresso sulla superficie delle cellule bersaglio. Poiché il riconoscimento TCR-pMHC avviene alla giunzione tra due cellule mobili, è stato a lungo sospettato di sperimentare forze meccaniche. Inoltre, questo ha portato al modello meccanosensore di attivazione TCR, che suggerisce che le forze TCR contribuiscono alla sua funzione 3,4. Per capire quando, dove e come le forze meccaniche contribuiscono alla funzione delle cellule T, è imperativo sviluppare strumenti per visualizzare le forze molecolari trasmesse dalle cellule T. Tradizionalmente, metodi come la microscopia della forza di trazione (TFM) e gli array di micropilastri sono usati per studiare le forze cellulari 5,6. Tuttavia, la sensibilità alla forza degli array di TFM e micropilastri è alla scala nanonewton (nN) e quindi è spesso insufficiente per studiare le forze molecolari di piconewton (pN) trasmesse dai recettori cellulari7. Per migliorare la forza e la risoluzione spaziale per il rilevamento, il nostro laboratorio ha aperto la strada allo sviluppo di sonde di tensione molecolare, che sono state inizialmente sintetizzate utilizzando polimeri di polietilenglicole (PEG)7. Le sonde di tensione molecolare sono costituite da una “molla” molecolare estensibile (PEG, proteina, DNA) affiancata da un fluoroforo e un quencher e sono ancorate su una superficie. Le forze applicate al terminale della sonda portano alla sua estensione, separando il fluoroforo e il quencher, e generando così un forte segnale di fluorescenza (Figura 1A)8,9,10.
Negli ultimi dieci anni abbiamo sviluppato una libreria di diverse classi di sonde di tensione molecolari con elementi a molla costituiti da acidi nucleici11, proteine10 e polimeri8. Tra questi, le sonde di tensione basate sul DNA forniscono il più alto rapporto segnale/rumore e la massima sensibilità alla forza, che è facilmente sintonizzata da pochi pN fino a ~20 pN11. Abbiamo utilizzato queste sonde di tensione del DNA in tempo reale per studiare le forze molecolari generate da molti diversi tipi di cellule, tra cui fibroblasti, cellule tumorali, piastrine e cellule immunitarie11,12,13. Questo manoscritto descriverà i protocolli per sintetizzare e assemblare sonde di tensione del DNA su una superficie per mappare le forze dei recettori molecolari con risoluzione della forza pN utilizzando un microscopio a fluorescenza convenzionale. Mentre l’attuale procedura include modifiche chimiche all’acido nucleico per introdurre il reporter fluorescente (Figura 1B), è importante notare che molte delle fasi di modifica e purificazione possono essere esternalizzate a società di sintesi del DNA personalizzate. Pertanto, la tecnologia delle sonde di tensione del DNA è facile e accessibile alle più ampie comunità di biologia cellulare e meccanobiologia.
In breve, per assemblare i sensori di tensione del DNA, una forcina di DNA viene ibridata con un filamento di ligando fluorescente su un braccio e un filamento di ancoraggio di quencher sull’altro braccio e quindi immobilizzato su un substrato di vetro (Figura 1C, tensione in tempo reale). In assenza di forza meccanica, la forcina viene chiusa e quindi la fluorescenza viene spenta. Tuttavia, quando la forza meccanica applicata è maggiore di F1/2 (la forza all’equilibrio che porta a una probabilità del 50% di dispiegarsi), la forcina si scioglie meccanicamente e viene generato un segnale fluorescente.
Basandoci sul sensore di tensione del DNA in tempo reale, descriviamo anche i protocolli per mappare le forze accumulate, che è particolarmente utile per studiare le interazioni tra i recettori sulle cellule immunitarie e il loro ligando naturale. Questo perché i recettori immunitari spesso mostrano legami di breve durata 3,14. Le forze accumulate vengono visualizzate utilizzando un filamento di “bloccaggio” che si lega preferenzialmente alle forcine di DNA aperte e consente la memorizzazione di segnali di fluorescenza associati a eventi di trazione meccanica (Figura 1C, tensione bloccata). Il filo di bloccaggio è progettato per legare un sito di legame criptico che viene esposto alla fusione indotta meccanicamente della forcina e bloccare la forcina nello stato aperto bloccando la ripiegatura della forcina, memorizzando così il segnale di tensione e generando una mappa di tensione accumulata. Inoltre, il filo di bloccaggio è progettato con un appiglio a otto nucleotidi, che consente una reazione di spostamento del filamento mediata dalla presa con il suo complemento completo, il filamento “sbloccante”. Con l’aggiunta del trefolo di sblocco, il filo di bloccaggio legato viene rimosso dal costrutto a forcina, cancellando il segnale di tensione memorizzato e ripristinando la forcina allo stato in tempo reale.
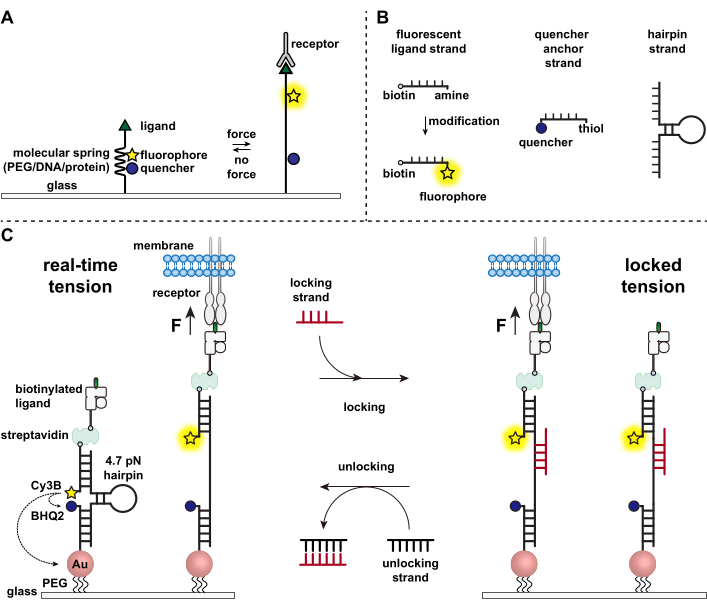
Figura 1: Schema delle sonde di tensione molecolari all’avanguardia . (A) Progettazione generale della sonda di tensione molecolare in tempo reale, (B) Filamenti per il costrutto della sonda di tensione basata sul DNA e (C) sonde di tensione basate sul DNA ingegnerizzate e loro commutazione tra stato in tempo reale e stato bloccato. Fare clic qui per visualizzare una versione ingrandita di questa figura.
Il protocollo principale consiste di quattro sezioni principali: preparazione degli oligonucleotidi, preparazione della superficie, imaging e analisi dei dati. Questo protocollo è stato dimostrato con successo dal nostro laboratorio e da altri in cellule T CD8+ OT-1 naïve e attivate, cellule CD4+ OT-II e ibridomi e può essere applicato per interrogare diversi recettori delle cellule immunitarie tra cui il recettore delle cellule T, il recettore della morte cellulare programmata (PD1) e le forze dell’antigene 1 associato alla funzione linfocitaria (LFA-1). Le cellule T CD8 + naïve OT-1 sono usate come linea cellulare di esempio in questo articolo.
