1. Preoperative Care
- Remove food from rat overnight prior to surgery.
- Induce anaesthesia in chamber with 4-5% isoflurane and O2 flow of 2 l/min.
- Shave abdomen from sternum to pelvis using electric razor.
- Place anaesthetised rat in supine position on isothermal heating pad.
- Apply eye ointment (Vitagel) before placing the rats’ snout in nosecone.
- Maintain anaesthesia with isoflurane concentration of 2-3% and O2 flow of 2 l/min.
- Disinfect skin with Betadine-Solution.
- Confirm depth of anaesthesia with forceps pinch between toes of hind leg.
- Administer 5.7 mg/kg Enrofloxacin intraperitoneally as perioperative antibiotic prophylaxis, and 1 mg/kg Flunixin for analgesia.
2. Median Laparotomy
- Perform midline incision using scalpel starting just below xyphoid process (Blade No. 10).
- Mobilise skin circumferentially from underlying abdominal muscles using Metzenbaum scissors.
- Open abdominal cavity.
- Install retractors to facilitate best possible exposure of the operation field.
3. Biliopancreatic and Alimentary Limb
- Identify where the duodenum or proximal jejunum passes under the colon.
- Transect small bowel about 10 cm aborally from here and ligate both ends of the gut (PDS 5-0).
- Place proximal stump of the two ends in left upper quadrant of abdomen as it will be later used to form the biliopancreatic limb of Roux-en-Y reconstruction.
- Place distal stump of the two ends in right upper quadrant of abdomen as it will be later used to form the alimentary limb of Roux-en-Y reconstruction.
4. Jejuno-Jejunostomy
- Identify caecum with ileocoecal valve and ileum.
- Follow ileum orally for approximately 25 cm. The Jejuno-Jejunostomy will be placed here as starting point of the common channel of Roux-en-Y reconstruction.
- Retrieve biliopancreatic limb from left upper quadrant of abdomen and position it next to common channel where you plan to perform Jejuno-Jejunostomy.
- Secure biliopancreatic limb and common channel with retention stitch (PDS 6-0).
- Incise both loops over approximate 10 mm by using micro scissors.
- Create Jejuno-Jejunostomy by performing side-to-side anastomosis using interrupted sutures (PDS 6-0).
- First complete dorsal side and then ventral side of anastomosis.
5. Gastric Pouch
- Identify gastro-oesophageal junction.
- Mobilise this area by dissecting gastro-hepatic and gastro-splenic ligaments using Metzenbaum scissors.
- Move left gastric artery and vagal fibres of left para-oesophageal bundle laterally to prevent major bleedings and vagal nerve damage when small gastric pouch is created.
- Expose gastro-oesophageal junction by placing cotton swab retro-oesophageally.
- Coagulate small vessels of frontal stomach by using commercially available cautery device – also to prevent bleedings.
- Transect stomach approximately 5 mm below gastro-oesophageal junction creating gastric pouch of a size of no more than 2-3% of original stomach size using delicate, curved scissors.
- Close gastric remnant (PDS 5-0).
6. Gastro-Jejunostomy
- Retrieve alimentary limb from right upper quadrant of abdomen and position it next to gastric pouch.
- Create Gastro-Jejunostomy by performing end-to-side anastomosis (PDS 7-0).
- First complete back side and then front side of anastomosis.
7. Abdominal Closure
- Reduce anaesthesia by reducing isoflurane concentration to 1.5%.
- Close muscle layer of abdominal wall using continuous sutures (PDS 4-0).
- Administer 100 μl of 0.3 mg/ml buprenorphine solution subcutaneously for analgesia.
- Further reduce isoflurane concentration down to 1%.
- Close the skin using interrupted sutures (Vicryl 4-0).
8. Postoperative Care
- Stop isoflurane and continue with O2.
- Administer 5 ml of warm saline for fluid replacement in three subcutaneous depots.
- Position rat under red light until full recovery.
- Return the rat to home cage.
9. Representative Results
Animals and housing
Male Wistar rats (Harlan Laboratories Inc., Blackthorn, UK; Elevage Janvier, Le-Genest-St. Isle, France) weighing between 350 and 500 g were individually housed under a 12 h /12 h light-dark cycle at a room temperature of 21±2 °C. Water and standard chow were available ad libitum, unless otherwise stated. All experiments were performed under a license issued by the Home Office, UK (PL70-6669) or approved by the Veterinary Office of the Canton Zurich, Switzerland. All rats were given one week of acclimatization before being randomized to gastric bypass or sham-operation. After surgery, rats received liquid diet for 3 days before access to normal chow was reinstalled.
Body weight
Data of our rat gastric bypass model are consistent with previous findings that gastric bypass surgery is effective to reduce body weight and especially to maintain body weight loss (Figure 2). Average pre-surgical body weight of rats used for gastric bypass and sham-operations was similar (sham: 433.4 ± 8.3 g vs. bypass: 420.7 ± 8.4 g, p= 0.28). Five days after surgery sham-operated controls weighed significantly more compared to gastric bypass rats (sham: 422.2 ± 8.3 g vs. bypass: 374.7 ± 7.6 g, p<0.001). On postoperative day 60, difference in body weight was almost 170 g (sham: 533.2 ± 8.1 g vs. bypass: 366.2 ± 10.8 g, p<0.001).
Food intake
Food intake followed similar patterns as body weight and was reduced in gastric bypass rats when compared to sham-operated ad libitum fed rats. Figure 3 shows the average daily food intake for both groups (postoperative day 1-60). Daily food intake was consistently lower after gastric bypass (sham: 29.9 ± 0.2 g vs. bypass: 25.7 ± 0.3 g, p<0.001).
Gut hormones
Blood from all rats was collected on the day of study termination 8,16. Animals had ad libitum food access the night before and were decapitated at the beginning of the light cycle on postoperative day 60. Blood was obtained, immediately centrifuged at 3000 rpm for 10 minutes at 4°C, and stored at -20°C until the samples were assayed in duplicate in a single run. PYY-like immunoreactivity was measured with a specific and sensitive radioimmunoassay, which measures, both the full length (PYY1-36) and the fragment (PYY3-36). GLP-1 was measured by established in-house radioimmunoassays17,18. Differences in food intake may be partly explained by increased postprandial plasma levels of peptide YY (PYY) and glucagon-like peptide 1 (GLP-1) as gastric bypass rats showed significantly higher levels for PYY (sham: 26 ± 2 pmol/L vs. bypass: 141 ± 14 pmol/L, p<0.001) and GLP-1 (sham: 40 ± 5 pmol/L vs. bypass: 215 ± 23 pmol/L, p<0.001; Figure 4).
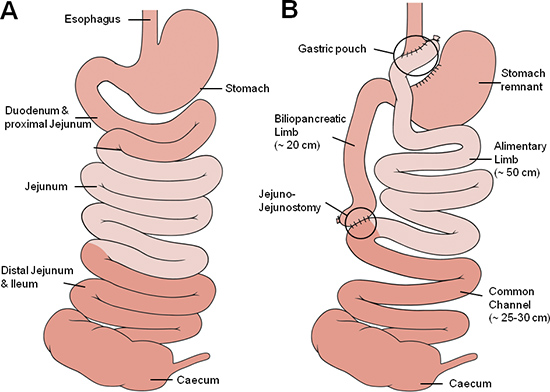
Figure 1. Gastric bypass anatomy. Schematic illustration of the small bowel anatomy before (A) and after (B) gastric bypass operation. The different shades of red approximately represent corresponding segments of the small bowel with the medium red representing the foregut (oesophagus, stomach duodenum and proximal jejunum), the light red representing the midgut (proximal and mid jejunum, proximal ileum) and the dark red representing the hindgut (ileum, caecum).
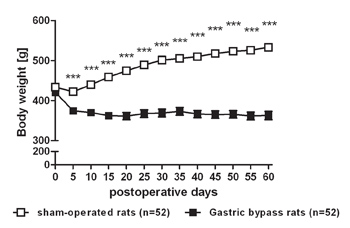
Figure 2. Body weight loss after gastric bypass surgery in rats. Body weight change for a representative group of rats after gastric bypass (- -) (n=52) and sham-operated rats (- -) (n=52) throughout an observation period of 60 days. Data were pooled from previous publications6,8-10 and are shown as mean values ± SEM (*** = p<0.001).
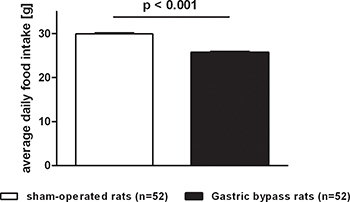
Figure 3. Average food intake after gastric bypass surgery in rats. Average daily food intake of a representative group of rats after gastric bypass (black, n=52) and sham-operated rats (white, n=52) throughout a postoperative period of 60 days. Data were pooled from previous publications6,8-10 and are shown as mean values ± SEM.
Figure 4. Postprandial PYY and GLP-1 serum levels after gastric bypass surgery in rats. Postprandial PYY and GLP-1 serum level for gastric bypass rats (black, n=18) and sham-operated rats (white, n=22). Data were pooled from previous publications8,16 and are shown as mean values ± SEM.
