Figure 1 depicts a screenshot of the customized software (NeuroLux Pro) developed in LabVIEW environment (National Instruments) used for simultaneous recording of neuronal activity and controlling light pulse parameters (i.e. pulse frequency, pulse width, stimulation period). The software program sends a command signal to a digital acquisition device that generates TTL pulses for control over the single diode laser. Additionally the software program displays and stores the recorded neuronal activity and delivered TTL signals. These signals are digitized through the acquisition device at a defined sample rate (e.g., 20 kHz). Recorded signals are saved as a two-dimensional double-precision array in a binary file.
Figure 2 shows the custom optrode consisting of a tungsten electrode attached to an optical fiber. An optical fiber is fixed to the electrode using thin suture thread at three points. If needed, super glue can be used to adhere them. However, avoid permanently fixing the optical fiber to the electrode because although the electrode may be reused several times, light transmission through the fiber will degrade with repeated use and should be cleaved and cleaned or replaced with every insertion into the brain.
Left panels of Figure 3 show fluorescent images of enhanced yellow fluorescent protein, indicating actual ChR2 and NpHR expression. These photographs show representative NpHR expression in rat dorsal subiculum (Figure 3A) and ChR2 expression in prelimbic cortex (Figure 3B). Viral expression was restricted mostly to the dorsal subiculum and prelimbic cortex (e.g. some fluorescence was observed in dentate gyrus (Figure 3A, left)). The tracks of the electrode (thin track, arrow head) and optical fiber (thick track, arrow) were also observed.
Left panel of Figure 4 shows that ChR2 induced temporally precise spiking in rat prelimbic cortex. Figure 4A is an example trace of ChR2-triggered action potentials in response to 20 Hz delivery of 10 msec blue light pulses to rat prelimbic cortex. Raster plot (Figure 4B) shows ChR2-induced activation in six representative neurons. All recorded neurons showed light-evoked spiking with perfect fidelity. In contrast to ChR2, NpHR rapidly and reversibly silenced spontaneous activity in vivo in rat prelimbic cortex (right panel of Figure 4). Figure 4C is an example trace showing that continuous 532 nm illumination (10 s) of the prelimbic cortex expressing NpHR under the CaMKllα promoter eliminates spontaneous single-unit activity in vivo. Note that the complete silencing of prelimbic pyramidal cell activity is time-locked to the 10 sec continuous light delivery. Lower raster plot (Figure 4D) shows NpHR-induced silencing in six representative neurons. Both in vivo recordings are typical of those obtained from adult Sprague-Dawley rats in which AAV-mediated delivery of microbial opsin genes occurred 18-21 days prior.
Figure 5 shows the result of repeated ChR2 stimulation of a rat prelimbic pyramidal neuron. Blue light pulse (10 msec) was delivered at 20 Hz for 5 sec (100 pulses total). Voltage traces of the light-evoked spiking were repeatedly acquired every 2 min during a 2 hr recording session (61 repetitions total). Even when using this repetitive stimulation protocol, ChR2 induced stable and robust spiking in response to the light delivery.
Figures 6 and 7 show the results of NpHR-induced photoinhibition and ChR2-driven photoactivation of rat subicular neuron activity. As is the case in prelimbic cortex, NpHR and ChR2 were able to mediate the light-induced time-locked inhibition and activation of the opsin-transduced subicular neurons with high reproducibility.
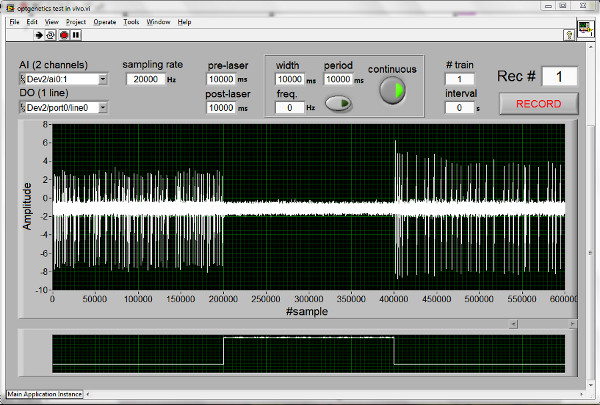
Figure 1. Screenshot of the NeuroLux Pro software interface for simultaneous light delivery and electrophysiological recording. Pictured trace shows NpHR induced silencing of spontaneous activity of rat prelimbic pyramidal cell in response to 10 sec of continuous 532 nm light delivery. Click here to view larger figure.
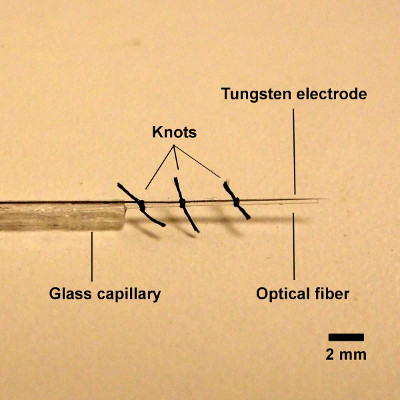
Figure 2. High power image of custom optrode. An optical fiber is inserted into glass capillary tube attached to a tungsten electrode and fixed to the electrode using suture thread. The center-to-center distance between the electrode tip and the fiber tip is approximately 500 μm.
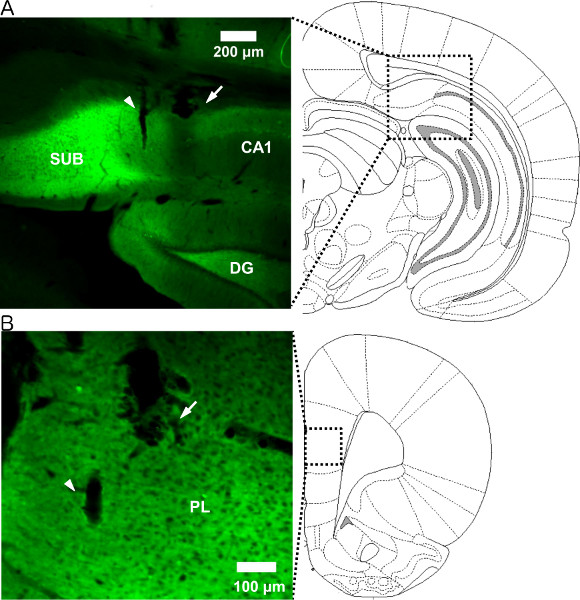
Figure 3. Viral expression and optrode placement. (Left) Photographs show representative NpHR expression in dorsal subiculum (A) and ChR2 expression in prelimbic cortex (B). (Right) Schematic that represents the location where each photograph was taken. The arrowhead and arrow in each photograph indicate the location of the tungsten electrode and optical fiber, respectively. SUB: subiculum, DG: dentate gyrus, PL: prelimbic cortex. Click here to view larger figure.
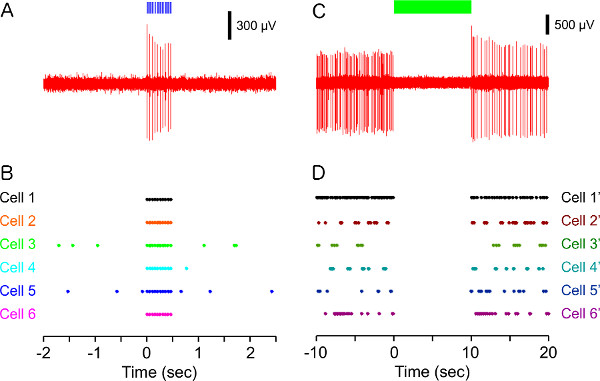
Figure 4. In vivo electrophysiological recordings from ChR2 or NpHR-transduced rat prelimbic cortex. (A) Example trace of ChR2-triggered action potentials in response to 20 Hz delivery of blue (473 nm) light pulses (10 msec, blue bar). (B) Raster plot showing ChR2-induced spiking in six representative neurons. Each unit activity is plotted as a dot. (C) Example trace of NpHR-induced suppression of spontaneous activity during continuous green (532 nm) light illumination (10 sec, green bar). (D) Raster plot showing NpHR-induced silencing in six representative neurons. Each unit activity is plotted as a dot. Click here to view larger figure.
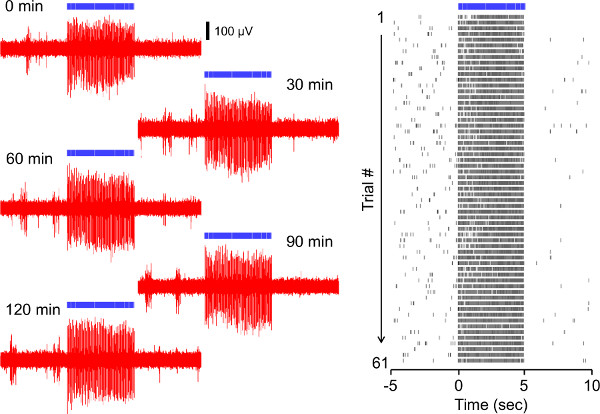
Figure 5. Repetitive 20 Hz ChR2-driven spiking of a rat prelimbic pyramidal neuron in vivo. (A) Voltage traces of the light-evoked spiking acquired at the time points 0, 30, 60, 90, 120 min after the beginning of the recordings. (B) Raster plot showing all 61 repetitions (2 min inter-trial interval, 120 min total) of the light-induced activation. Each unit activity is plotted as a dot. Click here to view larger figure.
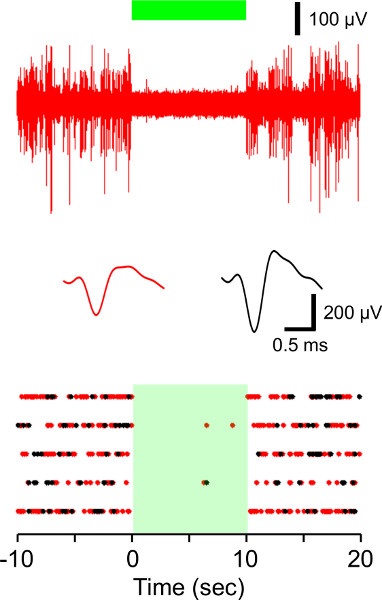
Figure 6. In vivo electrophysiological recordings from NpHR-transduced rat dorsal subiculum. (A) Example trace showing that 10 sec continuous 532 nm illumination (green bar) of the dorsal subiculum expressing NpHR eliminates spontaneous activity. (B) Average waveforms of the two recorded units from the above trace. The amplitude threshold was used for identifying two distinct neurons. (C) Raster plot showing five repetitions of NpHR-induced silencing of these units. Each unit activity is plotted as a dot.
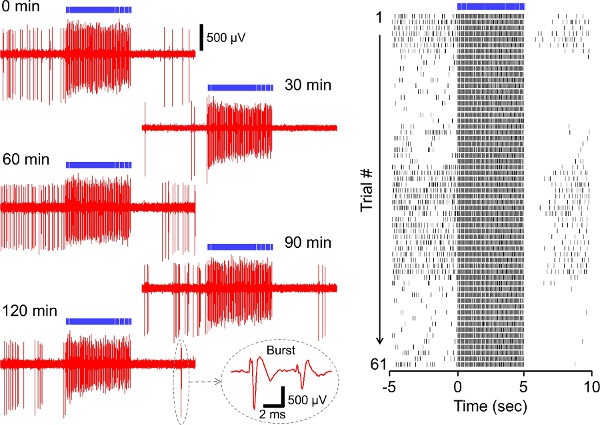
Figure 7. Repetitive 20 Hz CR2-driven spiking of a rat dorsal subicular neuron in vivo. (A) Voltage traces of the light-evoked spiking acquired at the time points 0, 30, 60, 90, 120 min after the beginning of the recordings. Inset: typical bursting activity of this cell. (B) Raster plot showing all 61 repetitions (2 min inter-trial interval, 120 min total) of the light-induced activation. Each unit activity is plotted as a dot. Click here to view larger figure.
