Renal epitel jontransport ligger bakom organism iono- och osmoregulation. Drosophila melanogaster Malpighian (renal) tubuli erbjuder en unik möjlighet att studera de molekylära mekanismerna för epitel jontransport. Detta beror på kombinationen av de kraftfulla genetik av Drosophila, ihopkopplade med tillgängligheten till sina njurtubuli till fysiologisk studie. The Ramsay-analysen, uppkallad efter den utredare som banat väg för tekniken 1, mäter vätska sekre priser från isolerade Malpighian tubuli, som grundades i Drosophila 1994 av Dow och kollegor 2. Detta banade väg för fortsatta studier med hjälp av Drosophila genetiska verktyg, såsom GAL4-UAS systemet 3,4, för att definiera cellspecifika signalvägar som reglerar vätskeutsöndring. Ett exempel innefattar kalciumsignalering som svar på ett peptidhormon 5, bland många andra 6,7.
ve_content "> En kombination av genteknik och klassisk fysiologisk studie har visat att urinproduktionen i farten sker genom utsöndring av en kaliumklorid rika vätska från huvud segment av tubuli. Detta sker genom parallell transepiteliala utsöndringen av katjoner, främst K + men också Na +, genom huvud cellen, och Cl -. sekre genom stelcellen 8-12 Möjligheten att separat mäta transepiteliala K + och Na + flöden tillåter en mer detaljerad beskrivning av transportmekanismer än mätning av vätskeutsöndring ensam. Till exempel i ostimulerade Drosophila tubuli, Na + / K + -ATPas-hämmare ouabain har ingen effekt på vätskeutsöndring 2, även om dess upptagning i huvud celler hämmas av den organiska anjontransportpolypeptiden hämmare taurokolat 13. dock Linton och O'Donnell visade att ouabain depolariserarden basolaterala membranpotentialen och ökar Na + flux 9. Som framgår av Representativa resultat, replikerade vi dessa resultat och visade att K + flödet är samtidigt minskade med 14; den ökade Na + flöde och minskad K + flux har motsatta effekter på vätskeutsöndring, vilket resulterar i någon nettoförändring av sekretion. Det finns alltså två resolutioner till "ouabain paradoxen", det vill säga den första observationen att ouabain har någon effekt på vätskeutsöndring i Drosophila tubuli. Först i stimulerade tubuli, är inte uppenbart att effekten av ouabain på vätskeutsöndring på grund av dess upptag av organic anion transporter 13; och andra, i ostimulerade tubuli, ouabain har motsatta effekter på transepitelial Na + och K + flöde, vilket resulterar i någon nettoförändring i vätskeutsöndring (se Representativa resultat och ref. 9). Därför primära roll Na + / K + -ATPas i ostimulerade tubuli är att sänka intracellulär Na + koncentration för att generera en positiv koncentrationsgradient för Na + -kopplade transportprocesser över basolaterala membranet. I själva verket, genom att separat mäta Na + och K + flussmedel, vi visade att tubuli saknar fly natrium-kalium-2-klorid cotransporter (NKCC) har minskat transepitelial K + flux, utan ytterligare minskning efter ouabain Dessutom, och ingen förändring i transepitelial Na * flux 14. Dessa upptäckter stödde vår slutsats att Na ^ in i cellen genom NKCC recirkuleras via Na * / K + -ATPas. I ett annat exempel, Ianowski et al. Konstaterade att sänka bad K + koncentration från 10 mM till 6 mM minskade transepiteliala K + flöde och ökad transepiteliala Na + flödet i tubuli från Rhodnius prolixus, utan nettoförändring i vätskeutsöndring <sup> 15. Särskiljande effekter på Na + flöde och K + flux över larv tubuli har också observerats i Drosophila tubuli som svar på varierande saltdieter 16 och två myggarter som svar på uppfödning salthalt 17.Den största utmaningen i mätningen av transepiteliala jonflöde i Ramsay analys preparatet är bestämningen av jonkoncentrationer inom utsöndrade vätskan. Denna utmaning har mötts med varierande lösningar, inklusive låga photometery 18, användning av radioaktiva joner 19 och elektronsond våglängd spektroskopi 20. Dessa tekniker kräver överföring av den utsöndrade vätskan sjunka till ett instrument för mätning av jonkoncentrationer. Eftersom volymen hos fluid utsöndras av ostimulerade Drosophila tubulus är liten, typiskt ~ 0,5 Nl / min, utgör detta en teknisk utmaning och dessutom införs fel om en del av den utsöndrade fluiden ärförloras vid överföring. Däremot möjliggör användning av jonspecifika elektroder mätningen av jonaktivitet (från vilken jonkoncentration kan beräknas) in situ. Den nuvarande protokollet anpassades från det som används av Maddrells och kollegor att mäta transepiteliala K + flöde över Rhodnius tubuli använder valinomycin som K + jonoforen 21, och även beskriver användningen av en 4-tert butylkalix [4] aren-tetraättiksyra tetraetylester baserade Na + specifik jon-specifik elektrod kännetecknas av Messerli et. al. 22. Jonspecifika elektroder har också använts för att mäta jonkoncentrationer i vätska som utsöndras från Malpighian tubuli i Ramsay analys i vuxen 9,23 och larver 16 Drosophila melanogaster, den nyzeeländska alpina Weta (Hemideina maori) 24 och i myggor 17.
Här beskriver vi i detalj användningen av Ramsay somsäga att mäta fluidsekrehastigheter i Malpighian tubuli från Drosophila melanogaster, liksom användning av jonspecifika elektroder för att bestämma koncentrationerna av K + och Na + inom den utsöndrade fluiden och därmed beräkningen av transepiteliala jonflöden. En översikt av analysen tillhandahålls i figur 1.
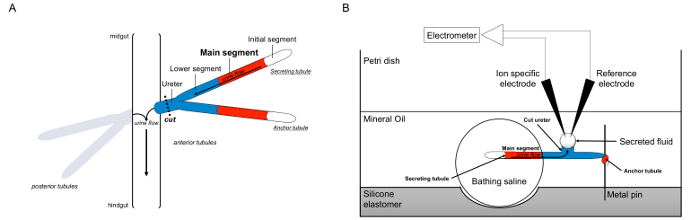
Figur 1. Skiss över Malpighian Sugrör och Ramsay analys med användning av Ion-specifika elektroder för att mäta jonkoncentrationer. Denna figur illustrerar inställningarna för den Ramsay analysen. (A) Varje fluga har fyra tubuli, ett par främre tubuli och ett par bakre kanalerna, som flyter i bukhålan omgiven av hemolymph. I varje par, de två små rören gå i urinledaren, som sedan tömmer urin vid korsningen av midgut och hindgut. Tubuli är blinda tills vidare. Urin genereras av fluid-utsöndrande huvudsegmentet (visat i rött), och strömmar mot urinledaren och ut i tarmen. Efter dissektion är tubuli paret skiljas från tarmen genom transecting urinledaren. (B) Paret av tubuli överförs sedan till en droppe av bad saltlösning inom en brunn av analys skålen. En av de två tubuli, hänvisas till här som den "ankare tubuli," lindas runt en metallstång och är inerta. Den andra tubuli är utsöndrar tubuli. Den initiala segmentet (som inte utsöndrar vätska) och huvudsegmentet av de utsöndrande tubulus förblir inom droppen av bad saltlösning. Joner och vatten skott från bad saltlösning och in i tubuli lumen i huvudsegmentet, och sedan röra sig mot urinledaren, som skulle ske in vivo. Det undre segmentet (blå) är utanför bad saltlösning och därmed inerta. Eftersom urinledaren kapas, framträder det utsöndrade fluiden som en droppe från den skurna änden av urinledaren. THan utsöndrade vätska droppe förstoras över tiden som sekre fortsätter och dess diameter mäts med hjälp av en okulär mikrometer. Ett lager av mineralolja förhindrar avdunstning av den utsöndrade vätskan. Referens- och jon specifika elektroder mäter jonkoncentrationen av den utsöndrade vätskan. Klicka här för att se en större version av denna siffra.
