Renale trasporto di ioni epiteliale sottende organismal iono- e osmoregolazione. La Drosophila melanogaster Malpighi (renale) tubulo offre un'opportunità senza precedenti per studiare i meccanismi molecolari del trasporto di ioni epiteliale. Ciò è dovuto alla combinazione delle potenti genetica di Drosophila, accoppiati con l'accessibilità dei suoi tubuli renali allo studio fisiologico. Il saggio Ramsay, dal nome del ricercatore che ha aperto la strada alla tecnica 1, misura i tassi di secrezione di fluidi da isolate tubuli malpighiani, ed è stato fondato nel Drosophila nel 1994 da Dow e colleghi 2. Questo ha spianato la strada a ulteriori studi utilizzando Drosophila strumenti genetici, come il sistema GAL4-UAS 3,4, per definire vie di segnalazione specifici delle cellule che regolano la secrezione di fluidi. Un esempio include segnalazione di calcio in risposta ad un ormone peptidico 5, tra molti altri 6,7.
ve_content "> una combinazione di tecniche genetiche e studio fisiologico classica ha dimostrato che la produzione di urina nella mosca avviene attraverso la secrezione di un potassio ricco cloruro fluido dal segmento principale del tubulo. Ciò avviene attraverso la secrezione transepiteliale parallelo di cationi, soprattutto K + ma anche Na +, attraverso la cella principale e Cl -. secrezione attraverso la cella stellate 8-12 La capacità di misurare separatamente transepiteliali K + e Na + flussi consente una caratterizzazione più dettagliata dei meccanismi di trasporto rispetto alla misurazione della secrezione di fluido alone. Ad esempio, in unstimulated tubuli Drosophila, Na + / K + -ATPasi inibitore ouabain non ha alcun effetto sulla secrezione di fluido 2, anche quando il suo assorbimento nelle cellule principali è inibito dal trasportatore anionico organico inibitore taurocolato 13. Tuttavia, Linton e O'Donnell ha mostrato che ouabain depolarizzail potenziale di membrana basolaterale, e aumenta Na + flusso 9. Come mostrato nelle Rappresentante dei risultati, abbiamo replicato questi risultati, e ha mostrato che K + flusso è contemporaneamente diminuito 14; l'aumento del flusso di Na + e diminuita K + flusso avere effetti opposti sulla secrezione di liquidi, con conseguente nessun cambiamento netto della secrezione. Così, ci sono due soluzioni a "paradosso ouabaina", cioè, l'osservazione iniziale che uabaina non ha alcun effetto sulla secrezione di fluido nel tubulo Drosophila:. Primo, in tubuli stimolato, l'effetto di ouabain sulla secrezione di fluido non è evidente a causa il suo assorbimento da parte del trasportatore anionico organico 13; e in secondo luogo, nei tubuli unstimulated, ouabain ha opposti effetti sulla transepiteliale Na + e K + flusso, con conseguente nessun variazione netta secrezione di fluidi (vedi Rappresentante dei risultati e rif. 9). Pertanto, il ruolo primario della Na + / K + -ATPase in tubuli non stimolati è quello di ridurre la concentrazione intracellulare di Na + per generare un gradiente di concentrazione favorevole per Na + -coupled processi di trasporto attraverso la membrana basolaterale. Infatti, misurando separatamente Na + e K + flussi, abbiamo dimostrato che tubuli manca la cotrasportatore fly sodio-potassio-2-cloruro (NKCC) sono diminuiti K transepiteliale + flusso, senza ulteriore calo dopo ouabaina aggiunta, e nessun cambiamento in transepiteliale Na + Flusso 14. Questi risultati supportati nostra conclusione che Na + di entrare nella cellula attraverso la NKCC viene riciclato attraverso il Na + / K + ATPasi. In un altro esempio, Ianowski et al. Ha osservato che l'abbassamento bagno K + concentrazione da 10 a 6 mm è diminuito transepiteliale K + flusso e aumento transepiteliale Na + flusso nel tubuli da Rhodnius prolixus, senza variazione netta secrezione di fluidi <sup> 15. Diversi effetti sulla Na + K + flusso e flusso attraverso tubuli larvali sono stati osservati anche nei tubuli Drosophila in risposta alle diverse diete sale 16 e in due specie di zanzare in risposta ad allevamento salinità 17.La sfida più grande nella misurazione del flusso transepiteliale di ioni nella preparazione dosaggio Ramsay è la determinazione delle concentrazioni di ioni all'interno del fluido secreto. Questa sfida è stata accolta con soluzioni diverse, tra cui la fiamma photometery 18, l'uso di ioni radioattivi 19, ed elettrone sonda lunghezza d'onda spettroscopia dispersiva 20. Queste tecniche richiedono trasferimento della goccia liquido secreto a uno strumento di misura della concentrazione di ioni. Poiché il volume di fluido secreto dalla non stimolato Drosophila tubulo è piccolo, tipicamente ~ 0,5 Nl / min, questo pone una sfida tecnica e introduce anche errore se parte del fluido è secretoperso al momento del trasferimento. Al contrario, l'uso di elettrodi ione-specifico permette la misura dell'attività ionica (da cui la concentrazione di ioni può essere calcolato) in situ. Il protocollo attuale è stato adattato da quella utilizzata da Maddrell e colleghi per misurare transepiteliale K + flusso attraverso il tubulo Rhodnius utilizzando valinomicina come il K + ionoforo 21, e descrive anche l'uso di un -butylcalix 4- ter [4] acido arene-tetraacetico tetraetile estere basato Na + -specifica elettrodi specifici ioni di litio caratterizzata da Messerli et. al. 22. Elettrodi Ion-specifici sono stati utilizzati anche per misurare le concentrazioni di ioni nel fluido secreto dalle tubuli malpighiani nel test Ramsay negli adulti 9,23 e larvale 16 Drosophila melanogaster, la Nuova Zelanda Alpine Weta (Hemideina maori) 24 e 17 zanzare.
Qui, descriviamo in dettaglio l'uso del Ramsaydire di misurare i tassi di fluido secrezione nei tubuli malpighiani da Drosophila melanogaster, nonché l'uso di elettrodi ione-specifico per determinare le concentrazioni di K + e Na + nel liquido secreto e quindi il calcolo dei flussi transepiteliale di ioni. Una panoramica del test è fornito in Figura 1.
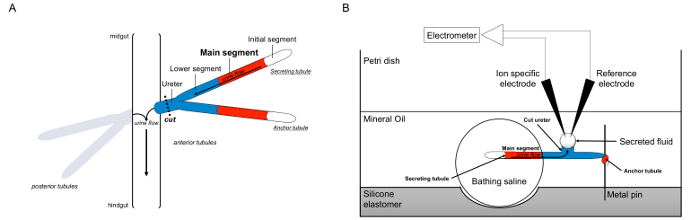
Figura 1. Schema del Malpighi tubuli e la Ramsay test con l'uso di ioni a specifici elettrodi per misurare concentrazioni di ioni. Questa figura illustra la configurazione per il saggio Ramsay. (A) Ogni mosca ha quattro tubuli, un paio di tubuli anteriori e una coppia di tubuli posteriori, che galleggiano nella cavità addominale circondato da emolinfa. In ogni coppia, i due si uniscono a tubuli l'uretere, che poi svuota l'urina all'incrocio tra midgut e hindgut. I tubuli sono ciechi a tempo indeterminato. L'urina viene generato attraverso il segmento principale del fluido-secernente (mostrato in rosso), e scorre verso l'uretere e fuori nell'intestino. Dopo la dissezione, la coppia tubulo è dissociata dall'intestino da sezionare l'uretere. (B) La coppia di tubuli viene poi trasferita in una goccia di soluzione salina in un bagno ben del piatto dosaggio. Uno dei due tubuli, qui definito il "ancoraggio tubulo," è avvolto attorno ad un perno di metallo ed è inerte. L'altro è il tubulo tubulo secernente. Il segmento iniziale (che non secernono liquido) e il segmento principale del tubulo secernono rimangono entro la goccia di soluzione salina balneazione. Ioni e spostare l'acqua dalla salina di balneazione e nel lume del tubulo del segmento principale, e poi si muovono verso l'uretere, come accadrebbe in vivo. Il segmento inferiore (blu) è fuori dalla salina di balneazione e quindi inerte. Poiché l'uretere è tagliato, il liquido secreto emerge come una gocciolina dall'estremità tagliata dell'uretere. Tegli secreto fluido gocciolina allarga nel tempo la secrezione continua, e il suo diametro viene misurato con un micrometro oculare. Uno strato di olio minerale impedisce l'evaporazione del fluido secreto. Gli elettrodi di riferimento specifici e ioni misurano la concentrazione di ioni di liquido secreto. Cliccate qui per vedere una versione più grande di questa figura.
