Rénale transport ionique épithéliale sous-tend organismal ionosphère et osmorégulation. Le Drosophila melanogaster Malpighian (rénale) tubule offre une occasion unique d'étudier les mécanismes moléculaires de transport d'ions épithéliale. Cela est dû à la combinaison de la génétique de la drosophile puissants, associé à l'accessibilité de ses tubules rénaux à l'étude physiologique. Le dosage Ramsay, nommé d'après le chercheur pionnier de la technique 1, mesure les taux de sécrétion de fluides à partir de tubes de Malpighi isolées, et a été établie chez la drosophile en 1994 par Dow et ses collègues 2. Cela a ouvert la voie à d'autres études utilisant la drosophile outils génétiques, telles que le système GAL4 UAS 3,4, pour définir les voies de signalisation cellulaires spécifiques régissant la sécrétion de fluide. Un exemple comprend la signalisation du calcium en réponse à une hormone peptidique 5, parmi beaucoup d'autres 6,7.
ve_content "> Une combinaison de techniques génétiques et étude physiologique classique a montré que la production d'urine chez la mouche se produit par la sécrétion d'un chlorure de potassium riche en fluide à partir du segment principal du tube. Ceci se produit par l'intermédiaire du transépithélial sécrétion parallèle de cations, principalement K + mais aussi Na +, à travers la cellule principale, et Cl -. sécrétion à travers la cellule stellaire 8-12 La capacité de mesurer séparément transépithéliaux flux K + et Na + permet une caractérisation plus détaillée des mécanismes de transport que la mesure de la sécrétion de fluide seul. Par exemple, dans les tubules Drosophila non stimulées, la Na + / K + -ATPase ouabaïne n'a aucun effet sur la sécrétion de fluide 2, même lorsque son absorption dans les cellules principales est inhibée par l'inhibiteur de transport anionique organique taurocholate 13. Cependant, Linton et O'Donnell a montré que l'ouabaïne dépolarisele potentiel de la membrane basolatérale, Na + et augmente le flux 9. Comme le montrent les résultats représentatifs, nous avons reproduit ces résultats, et montré que flux de K + est concomitante diminué de 14; l'augmentation du flux de Na + et K + a diminué flux des effets opposés sur la sécrétion de fluide, de sorte qu'aucun changement net de la sécrétion. Ainsi, il ya deux résolutions au «paradoxe ouabaïne», à savoir l'observation initiale que ouabaïne n'a aucun effet sur la sécrétion de fluide dans le tubule Drosophila:. La première, dans les tubules stimulés, l'effet de la ouabaïne sur la sécrétion de fluide est pas évident en raison de son absorption par le système de transport anionique organique 13; et la seconde, dans les tubules non stimulées, ouabaïne a des effets sur transepithelial Na + et K + flux opposés, entraînant aucun changement net de la sécrétion de fluide (voir résultats représentatifs et réf. 9). Par conséquent, le rôle principal de la Na + / K + ATPase dans les tubules non stimulées est de réduire la concentration intracellulaire de Na + à générer un gradient de concentration favorable pour Na + -coupled processus de transport à travers la membrane basolatérale. En effet, en mesurant séparément Na + et K + flux, nous avons démontré que les tubules manque la volée sodium-potassium-2-chlorure cotransporteur (NKCC) ont diminué K transepithelial + flux, sans diminution supplémentaire après addition ouabaïne, et aucun changement dans transepithelial Na + flux 14. Ces résultats ont appuyé notre conclusion que Na + entrant dans la cellule par l'intermédiaire du NKCC est recyclée à travers la Na + / K + -ATPase. Dans un autre exemple, Ianowski et al. Observe que l'abaissement de bain concentration de K + de 10 à 6 mm diminué transepithelial flux de K + et une augmentation transepithelial Na + flux dans les tubules de Rhodnius prolixus, avec aucun changement net de la sécrétion de fluide <sup> 15. Les effets différentiels sur Na + et K + flux flux à travers les tubules larvaires ont également été observés dans les tubules de drosophile en réponse à divers régimes de sel 16 et dans deux espèces de moustiques en réponse à l'élevage de la salinité 17.La plus grande difficulté dans la mesure de flux transépithélial d'ions dans la préparation d'essai Ramsay est la détermination des concentrations des ions dans le fluide sécrété. Ce défi a été relevé avec différentes solutions, y compris la flamme photometery 18, l'utilisation d'ions radioactifs 19 et sonde électronique longueur d'onde de la spectroscopie à dispersion 20. Ces techniques nécessitent le transfert de la goutte de fluide sécrété à un instrument de mesure des concentrations d'ions. Etant donné que le volume de fluide sécrété par les tubules Drosophila non stimulé est petite, typiquement environ 0,5 nl / min, ce qui pose un défi technique et introduit également erreur si une partie du fluide est sécrétéperdu au moment du transfert. En revanche, l'utilisation d'électrodes spécifiques des ions permet la mesure de l'activité ionique (à partir de laquelle la concentration en ions peut être calculée) in situ. Le protocole actuel a été adaptée à partir de celle utilisée par les Maddrell et collègues pour mesurer transépithélial flux de K + à travers le tube Rhodnius utilisant la valinomycine comme K + ionophore 21, et décrit également l'utilisation d'un -butylcalix 4- tert [4] d'acides arène-tétraacétique Na tétraéthyle à base d'ester + électrode spécifique-ion spécifique de caractérisé par Messerli et. . 22 al. Électrodes ions spécifiques ont également été utilisés pour mesurer les concentrations d'ions dans le liquide sécrété par les tubules de Malpighi dans le dosage Ramsay chez l'adulte et larvaire 9,23 16 Drosophila melanogaster, la Nouvelle-Zélande Alpine Weta (Hemideina maori) 24 et 17 chez les moustiques.
Ici, nous décrivons en détail l'utilisation de la Ramsay commedire pour mesurer les taux de sécrétion de fluide dans les tubes de Malpighi de Drosophila melanogaster, ainsi que l'utilisation d'électrodes spécifiques des ions afin de déterminer les concentrations de K + et Na + dans le fluide sécrété, et donc le calcul des flux transépithélial d'ions. Une vue d'ensemble de l'essai est fourni à la figure 1.
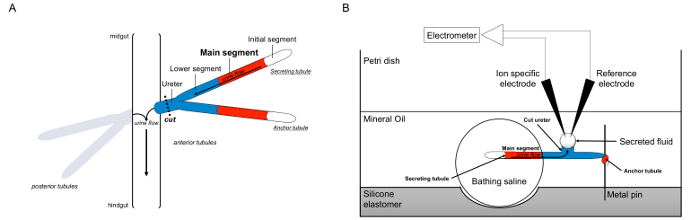
Figure 1. Schéma de l'Malpighian Pailles et le test Ramsay avec utilisation de l'ion-spécifiques électrodes pour mesurer les concentrations d'ions. Cette figure illustre la configuration pour le dosage Ramsay. (A) Pour chaque volée comporte quatre tubes, une paire de tubules antérieure et une paire de tubules postérieures, qui flottent dans la cavité abdominale entouré d'hémolymphe. Dans chaque paire, les deux tubes se rejoignent à l'uretère, qui se jette ensuite l'urine à la jonction de l'intestin moyen et hindgut. Les tubules sont aveugles-clos. L'urine est généré par le segment principal de fluide sécrétant (en rouge), et coule vers l'uretère et sortir dans l'intestin. Après dissection, la paire de tubule est dissocié à partir de l'intestin par transection l'uretère. (B) La paire de tubes est ensuite transféré dans une gouttelette de solution saline dans un bain bien du plat de dosage. L'un des deux tubes, appelé ici le "tube d'ancrage», est enroulé autour d'une tige de métal et est inerte. L'autre tube est le tube de sécrétion. Le segment initial (qui ne sécrètent pas fluide) et le segment principal de la tubule sécrétant restent dans la gouttelette de solution saline de baignade. Ions et mouvement de l'eau de la solution saline de bain et dans la lumière des tubules du segment principal, puis se déplacent vers l'uretère, comme cela se produirait in vivo. Le segment inférieur (bleu) est en dehors de la solution saline de baignade et donc inerte. Depuis l'uretère est coupé, le fluide sécrété apparaît comme une gouttelette à partir de la fin de l'uretère de coupe. Til agrandit gouttelette de fluide sécrété dans le temps comme la sécrétion continue, et son diamètre est mesuré en utilisant un micromètre oculaire. Une couche d'huile minérale empêche l'évaporation du fluide sécrété. Les électrodes spécifiques référence et d'ions de mesurer la concentration en ions du fluide sécrété. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
