תחבורת יון אפיתל כליות בבסיס iono- האורגניזם וויסות האוסמוטי. אבובית דרוזופילה melanogaster Malpighian (כליות) מציעה הזדמנות שאין כמותו כדי לחקור את המנגנונים המולקולריים של תחבורת יון אפיתל. זאת בשל השילוב של הגנטיקה החזקה של דרוזופילה, יחד עם הנגישות של tubules הכליות שלה למחקר פיסיולוגי. Assay ראמזי, על שמו של החוקר שהיה חלוץ טכניקת 1, מודד שיעורי הפרשת נוזל מצינוריות Malpighian מבודדות, והוקם בשנת 1994 על ידי תסיסנית דאו ועמיתים 2. זה סלל את הדרך למחקרים נוספים באמצעות תסיסנית כלים גנטיים, כגון מערכת GAL4-כטב"מ 3,4, להגדיר מסלולי איתות תא ספציפי המסדירים את הפרשת נוזלים. דוגמא כוללת איתות סידן בתגובה להורמון פפטיד 5, בקרב רבים אחרים 6,7.
ve_content "> שילוב של טכניקות גנטיות ומחקר פיסיולוגי קלאסי הוכיח כי דור שתן בזבוב מתרחש באמצעות ההפרשה של נוזל כלוריד-עשיר אשלגן מהמגזר העיקרי של אבובית. זו מתרחשת באמצעות הפרשת transepithelial המקבילה של קטיונים, בעיקר K + אלא גם Na +, דרך התא העיקרי, וCl -. ההפרשה דרך תא stellate 8-12 היכולת למדוד בנפרד והנתיבים transepithelial K + וNa + מאפשרת אפיון מפורט יותר של מנגנוני תחבורה מאשר המדידה של הפרשת נוזל יש לבד. לדוגמא, בtubules תסיסנית unstimulated, + Na / K + -ATPase מעכב ouabain אין כל השפעה על הפרשת נוזל 2, גם כאשר ספיגתו לתוך תאים עיקריים היא מעוכבת על ידי taurocholate מעכב טרנספורטר אניון האורגני 13. עם זאת, לינטון ו אודונל הראה כי ouabain depolarizesפוטנציאל הממברנה basolateral, ומגביר Na + שטף 9. כפי שניתן לראות בנציגי התוצאות, אנו משוכפלים ממצאים אלה, והראו כי K + שטף הוא במקביל ירד 14; שטף Na + המוגבר וירידת K + שטף יש השפעות מנוגדות על הפרשת נוזל, וכתוצאה מכך אין שינוי נטו בהפרשה. לפיכך, יש שתי החלטות ל" פרדוקס ouabain, "כלומר, ההתבוננות הראשונית שouabain אין כל השפעה על הפרשת נוזלים באבובית תסיסנית:. ראשונה, בtubules המגורה, ההשפעה של ouabain על הפרשת נוזל אינה נראית לעין עקב הספיגה שלה על ידי טרנספורטר אניון האורגני 13; ושנייה, בtubules unstimulated, ouabain יש השפעות מנוגדות על transepithelial Na + K + ושטף, וכתוצאה מכך אין שינוי נטו בהפרשת נוזל (ראה תוצאות ונ"צ נציג. 9). לכן, התפקיד העיקרי של + Na / K + -ATPASE בtubules unstimulated הוא להוריד תאיים ריכוז Na + ליצור שיפוע ריכוז נוח לNa + -coupled תהליכי הובלה על פני קרום basolateral. ואכן, על ידי מדידה בנפרד Na + K + ונתיבים, שהוכחנו כי tubules חסר cotransporter נתרן, אשלגן-2-כלוריד זבוב (NKCC) ירד K + שטף transepithelial, ללא ירידה נוספת לאחר תוספת ouabain, ולא חלו שינוי בtransepithelial Na + שטף 14. ממצאים אלה נתמכים מסקנתנו כי Na + כניסה לתא דרך NKCC ממוחזרת דרך Na + / K + -ATPase. בדוגמא אחרת, Ianowski נצפה et al. כי הורדת K אמבטיה + ריכוז מ -10 מ"מ ועד 6 מ"מ ירד K transepithelial + שטף וtransepithelial המוגבר שטף Na + בצינוריות מprolixus Rhodnius, ללא שינוי נטו בהפרשת נוזל <sup> 15. אפקטי ההפרש בשטף Na + K + ושטף על פני tubules זחל גם נצפו בtubules דרוזופילה בתגובה לדיאטות מלח שונה 16 ובשני מיני יתושים בתגובה לגידול מליחות 17.האתגר הגדול ביותר במדידת שטף יון transepithelial בהכנת assay רמזי הוא קביעת ריכוזי יון בתוך הנוזל המופרש. אתגר זה כבר נפגש עם פתרונות שונים, כולל photometery להבה 18, שימוש ביונים רדיואקטיביים 19, וגל אלקטרון הבדיקה ספקטרוסקופיה נפיצה 20. טכניקות אלה דורשות העברה של ירידת הנוזל המופרשת למכשיר למדידת ריכוזי יון. מאז את נפח הנוזל המופרש על ידי אבובית תסיסנית unstimulated הוא קטן, בדרך כלל ~ 0.5 NL / דקה, זה מציב אתגר טכני וגם מציגה שגיאה אם חלק מהנוזל המופרש הואאיבד על העברה. לעומת זאת, השימוש באלקטרודות יון ספציפי מאפשרת המדידה של פעילות יון (שממנו ניתן לחשב ריכוז יון) באתרו. הפרוטוקול הנוכחי הותאם מזה בשימוש על ידי Maddrell ועמיתים למדידת K transepithelial + שטף על פני אבובית Rhodnius באמצעות valinomycin כK + ionophore 21, ומתאר גם את השימוש ב-butylcalix 4 טרט [4] חומצת arene-tetraacetic Na מבוסס אסתר tetraethyl + אלקטרודה יון ספציפי -specific המאופיינת Messerli et. אל. 22. אלקטרודות יון ספציפיות יש גם שימשו למדידת ריכוזי יון בנוזל המופרש על ידי צינוריות Malpighian בassay רמזי ב9,23 מבוגרים ו -16 זחל תסיסנית melanogaster, ניו זילנד Alpine Weta (Hemideina מאורי) 24 וביתושים 17.
כאן, אנו מתארים בפירוט את השימוש ברמזים כאומר למדוד שיעורי הפרשת נוזל בtubules Malpighian מדרוזופילה melanogaster, כמו גם שימוש באלקטרודות יון ספציפי כדי לקבוע את הריכוזים של K + וNa + בתוך הנוזל המופרש ולכן החישוב של נתיבי יון transepithelial. סקירה של assay מסופקת באיור 1.
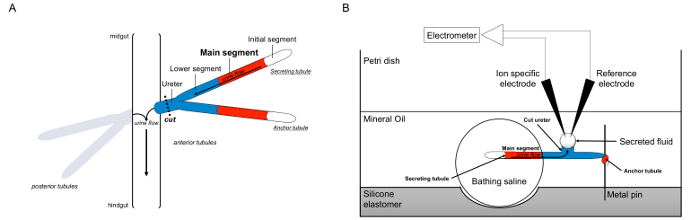
איור 1. סכמטי של קשיות Malpighian וAssay רמזי עם שימוש ביון ספציפי אלקטרודות למדידת ריכוזי יון. נתון זה ממחיש את ההתקנה לassay רמזי. (א) לכל זבוב ארבעה tubules, זוג הצינוריות קדמית וזוג הצינוריות אחורית, לצוף שבחלל הבטן המוקף hemolymph. בכל זוג, שתי הצינוריות להצטרף בשופכן, אשר לאחר מכן מרוקן את השתן בצומת של midgut וhindguלא. Tubules הם הסתיים עיוור. שתן נוצר על ידי המגזר העיקרי מפרישי הנוזל (באדום), וזורם לכיוון השופכן והחוצה אל הבטן. לאחר נתיחה, זוג אבובית הוא ניתק מהבטן על ידי חוצה את השופכן. (ב) זוג הצינוריות מועבר לאחר מכן לטיפה של ים מלוח בתוך היטב של צלחת assay. אחד משתי הצינוריות, המכונה כאן "אבובית העוגן," עטוף סביב סיכת מתכת והוא אינרטי. אבובית האחרות היא אבובית הסתרתו. הקטע הראשוני (שאינו מפריש נוזל) והקטע העיקרי של אבובית מפרישות להישאר בתוך הטיפה של ים מלוח. יונים ומהלך מים מתמיסת מלח הים ולתוך לום אבובית של המגזר העיקרי, ולאחר מכן לנוע לכיוון השופכן, כפי שהיו קורים בvivo. הקטע התחתון (הכחולה) הוא מחוץ למלוח רחצה ולכן אדיש. מאז שופכן הוא לחתוך, הנוזל המופרש מתגלה כטיפה מסוף החתך של השופכן. Tהוא מופרש טיפת נוזל מגדילה לאורך הזמן כהפרשה ממשיכה, וקוטרה נמדד באמצעות מיקרומטר עיני. שכבה של שמן מינרלים מונעת אידוי של הנוזל המופרש. אלקטרודות ספציפיות ההתייחסות והיון למדוד את ריכוז היון של הנוזל המופרש. אנא לחץ כאן כדי לצפות בגרסה גדולה יותר של דמות זו.
